
陈波 1,2陈巍 3,4张静 4袁逖飞 5
(1北京爱生科贸有限公司科仪部, 北京 100086) (2杭州师范大学认知与脑疾病研究中心, 杭州 310015) (3绍兴文理学院心理学系, 绍兴 312000) (4浙江大学语言与认知研究中心, 杭州 310028) (5南京师范大学心理学院, 南京 210097)
摘要
镜像神经元被认为是改变认知神经科学的重要发现,它的发现为迄今为止许多行为实验难以检验的心智或认知现象提供了一个统一的研究视角。然而,镜像神经元在社会认知活动中的重要作用却引起了研究者的激烈争论。系统回顾镜像神经元研究中的争议,可以尝试以争论焦点出现的逻辑关系和性质为依据将其划分为两类:一类是关于镜像 “内涵”的争议,主要包括镜像神经元是否存在及其起源的争议。另一类则主要围绕镜像 “外延”的争议, 即有关镜像神经元系统和镜像机制的争议。未来的研究必须摒弃将镜像神经元视为 “认知科学圣杯 ”的神经还原主义倾向,将之视为一个由多个子系统组成的开放的复杂巨系统,并将焦点转向关注镜像机制。
关键词
镜像神经元; 镜像内涵; 镜像外延; 镜像神经元系统; 镜像机制
分类号 B845
1 前言
意大利帕尔马大学的 di Pellegrino, Fadiga, Fogassi, Gallese和 Rizzolatti (1992)利用单细胞记录技术在恒河猴的运动前区皮层下部边缘位置 (F5区, Brodmann 6区腹侧边缘 )首次发现了一些具有特殊属性的神经元 , 它们在恒河猴自身执行目标导向 (goal-directed)手部动作或者观察同类执行相似动作时都被激活。由于这些神经元能像镜子一样映射其他个体的活动 , Gallese, Fadiga, Fogassi和 Rizzolatti (1996)和 Rizzolatti, Fadiga, Gallese和 Fogassi (1996)在进一步的单细胞记录研究中将其称为“镜像神经元” (mirror neurons)。镜像神经元的发现所带来的影响无疑是革命性的, 它们将研究者的目光从对知觉 –行为的探索转向了寻找人类镜像神经元活动的跨学科研究,这对个体的动作模仿学习、共情、语言习得和意图理解等社会认知活动的展开也具有极为重要的意义。镜像神经元甚至被描述为“心理科学的DNA”或 “塑造文明的神经元 ” (Ramachandran, 2011)。然而, 虽然在镜像神经元发现至今的余年内 , 相关的研究报告一直以几何级数递增 , 但研究者却很难从这些纷繁的研究成果中听到和谐的声音 , 更多的是无休止的激烈争论。这些争论有的来自严谨的实验室实验(Eshuis, Coventry, & ulchanova, 2009; Lingnau, Gesierich, & Caramazza,2009), 有的则来自理论性的反思 (Hickok, 2009, 2013; Gallese, Gernsbacher, Heyes, Hickok, & Iacoboni, 2011; Kosonogov, 2012; Casile, 2013;Caramazza, Anzelotti, Strnad, & Lingnau, 2014)、哲学思辩 (Borg, 2007, 2012; Uithol, Van Rooij, Bekkering, & Haselager, 2011)或者对当前研究的元分析 (Molenberghs, Cunnington, Mattingley, 2009,2012)。由此带来的影响直接质疑了 20年来基于镜像神经元及其系统提出的种种假设 , 包括动作目的的编码与模仿 (Cook & Bird, 2013; Caggianocomet al., 2013), 以及自闭症的 “碎镜理论 ” (Fan, Decety, Yang, Liu, & Cheng, 2010; Hamilton, 2013)。因此,系统澄清其中的争议将有助于研究者正确认识镜像神经元的功能,从而把握住镜像神经元研究未来发展的脉搏。由于镜像神经元在动作理解 (action understanding)与模仿 (imitation)上的作用是有关其功能最初也是最保守的假设 (Rizzolatti & Craighero, 2004; Hickok & Hauser, 2010; Rizzolatti & Sinigaglia, 2010; Frith & Frith, 2012), 对镜像神经元在动作理解上作用的质疑将直接构成对相关理论的瓦解 (Hickok, 2009; Kosonogov, 2012)。因此, 梳理该领域中存在的争议将成为所有相关讨论的基础。在本文中,作者尝试以这些争论焦点出现的逻辑关系和性质为依据将其划分为两类:一类是关于镜像 “内涵”的争议,主要包括镜像神经元是否存在及其起源的争议。另一类则主要围绕镜像“外延”的争议, 即有关镜像神经元系统 (mirror neuron system)和镜像机制(mirror mechanism)的争议。
2 关于镜像“内涵”的争议
di Pellegrino等(1992)在恒河猴的 F5区发现镜像神经元之后,提出了另一个十分有趣且非常值得思考的问题,即 F5区在人脑中的同源体大部分在 Broca区, 这是否是巧合? Gallese等(1996)和 Rizzolatti, Fadiga, Gallese等(1996)在利用单细胞记录技术绘制恒河猴镜像神经元脑区分布时同时注意到了这个问题,F5区在人脑同源体中是否真的也存在镜像神经元? Rizzolatti, Fadiga, Matelli等(1996)等利用 PET技术取得了一些收敛性的证据 (converging evidence), 并认为在人脑的颞中回 (middle temporal gyrus, Brodmann21区)、颞上沟 (superior temporal sulcus, STS)和额下回尾部(inferior frontal gyrus, IFG, Brodmann45区)存在具有镜像属性 (mirror properties)的神经元。由此拉开了关于人脑中镜像神经元是否存在及其起源,镜像神经元系统与镜像机制等问题争论的大幕。
目前, 由于研究对象和技术的局限性, 研究者不可能随意在人脑疑似镜像神经区域进行单个神经元神经电生理活动的研究。人脑中是否存在镜像神经元的确凿证据一直无法获得。幸运的是,当前在这一问题上已经取得了一些突破性的进展, 越来越多的证据指向人脑中存在镜像神经元这一假设。此外,同样引人关注的还有镜像神经元的产生机制, 对该问题的争议还未得到彻底解决。
2.1人脑中镜像神经元的存在
在很长一段时间里,关于人脑中是否存在镜像神经元的争论表面上可能更多地集中在其研究结果的间接性上。恒河猴 F5区混合存在着空间位置非常接近的视觉属性的、运动属性的、视觉–运动属性和听觉 –视觉属性的镜像神经元 (Kohler et al., 2002; Keysers et al., 2003)。人脑皮层中可能也存在相似的神经元 (Shmuelof & Zohary, 2005; Gazzola, Aziz-Zadeh, & Keysers, 2006; Keysers, 2009; Mukamel, Ekstrom, Kaplan, Iacoboni, & Fried, 2010)。由于当下广泛采用的 PET和 fMRI脑成像技术只能通过被试在有效执行动作和有效观察动作过程中所激活的脑区存在着空间重叠部分来证明镜像神经元的存在,但这些成像技术的分辨率使它们无法确定在有效执行动作和有效观察动作过程中都有激活的三维像素点中的神经元是相同的(Chong, Cunnington, Williams, Kanwisher, & Mattingley, 2008; Dinstein, Thomas, Behrmann, & Heeger, 2008; Dinstein, 2008; Keysers & Gazzola, 2009a, 2009b)。
造成这种争论的实质是:研究中的镜像神经元缺少操作性定义,或没有准确的操作性定义。那么, 究竟何为镜像神经元? Rizzolatti和 Craighero (2004)在迄今为止该领域具有广泛影响力的一篇综述中明确给出了操作性定义: (1)镜像神经元本身是一群特殊的运动神经元 (motor neuron)。即, 它首先应在个体执行动作时放电 (motor discharge); (2)除了在运动时放电,镜像神经元的活动还必须具备感觉 –运动的对应关系 (sensory-motor association)。例如, 几乎所有镜像神经元在对感觉刺激(包括动作的视觉刺激及与动作相关的听觉刺激)的反应与其编码的运动反应方面呈现出一致性。即,不仅仅是在自身执行某一动作时放电,而且在观察到他人做类似动作时也放电 (Glenberg, 2011)。这就是“镜像” (mirror)的意义。这两个特征将许多仅具备镜像属性的神经元排除了出去。例如, Hutchison, Davis, Lozano, Tasker和 Dostrovxky (1999)等曾以接受扣带回切断术 (Cingulotomy)的患者为研究对象,发现在人类大脑的扣带皮层 (cingulate cortex)上中存在一类与疼痛有关的神经元, 它使得人类的共情体验会出现一种 “镜像现象” (mirror phenomenon)。这种神经元不仅对作用于患者有害的机械刺激(例如, 针刺手指)产生反应, 也会对患者观察作用于实验者的相同刺激产生反应。不过,虽然这类神经元与镜像神经元在功能上极其类似,但它们是一种感觉神经元 (sensory neurons), 不具备运动属性,也就不能被归入镜像神经元。
以此作为标准不难发现,在当前的研究中存在对“镜像神经元 ”这一术语的滥用。一方面, 一些研究者缩小了镜像的内涵,只承认经典镜像神经元系统(classical mirror neuron system)中(额下回的后部、顶叶皮层 )的神经元统为镜像神经元。另一方面,另一些研究者则任意扩大了镜像的内涵,将只具备镜像属性而不具备运动属性的神经元也纳入镜像神经元的范畴 (Brown & Brüne, 2012)。这导致研究者无法确定在实验中观察到的被激活的神经元就是其所要研究的镜像神经元。
Gallese等(1996)曾采用单细胞记录技术在恒河猴 F5区植入了 532个电极,其中仅有 92个被记录的神经元被认为是镜像神经元。换言之,F5区只有很小一部分是镜像神经元。此外,他们依据视觉 (有效观察动作 ).运动(有效执行动作 )一致性将镜像神经元划分为了 3个子集, 即严格一致性 (strictly congruent)镜像神经元、宽泛一致性(broadly congruent)镜像神经元、不一致 (no congruent)镜像神经元。这三类镜像神经元分别占所记录到的镜像神经元总数的 31.5%、60.9%和 7.6%。
事实上这种根据神经元的反应作出的分类并不严谨,特别是弱化了对了镜像神经元的操作性定义。 Rizzolatti和 Craighero (2004)在提及这种分类时, 只提到了严格一致性镜像神经元和宽泛一致性镜像神经元。这可能是因为很多研究者已经开始意识到这种分类存在不合理性。此外,Rizzolatti和 Luppino (2001)将恒河猴 F5区的视觉运动神经元分为两类,即规范性神经元 (canonical neurons)和镜像神经元。规范性神经元对某一对象表征做出反应(例如, 观察到一个可被抓取的苹果), 而镜像神经元则对对象的意向性行为做出反应(例如, 观察到一个抓取苹果的动作 )。虽然这些分类有利于我们对研究中的镜像神经元进行操作性定义,但囿于现有脑成像技术分辨率的局限,它们仍然不能帮助我们确定在有效执行动作和有效观察动作过程中所激活的是相同的神经元。
为了改变这种尴尬的局面,一些研究者呼吁从神经元对动作的选择性上来完成对镜像神经元的操作性定义。于是 fMRI-A (Grill-Spector & Malach, 2001)的改进范式,即重复抑制 (repetition suppression)范式或适应 (adaptation)范式被引入研究, 它也被认为是目前识别镜像神经元的有效方法(Shmuelof & Zohary, 2005; Hamilton & Grafton, 2006; Dinstein, Hasson, Rubin, & Heeger, 2007; Dinstein et al., 2008; Dinstein, 2008)。这种方法的逻辑是:对于假定存在镜像神经元的特定皮层,重复动作刺激将导致该区域在单一重复抑制 (unimodal repetition suppression)和交互重复抑制 (cross-modal repetition suppression)下均出现适应现象(前者是指在先观察后执行或先执行后观察的情况下出现适应现象,后者则是指两种执行顺序的情况下都出现适应现象)。
遗憾的是,Dinstein等(2007)、Chong等(2008)和 Lingnau等(2009)尽管在单一模式下观察到了适应现象,但均未能在交互模式下观察到双向的适应现象。换言之,他们的研究并没有成功地分离出镜像神经元。 Kilner, Neal, Weiskopf, Friston和 Frith (2009)在他们的基础上改进了实验中的动作刺激和动作任务,使这些动作更具有 “意义”和更“敏感”, 并最终获得成功,他们首次在交互重复抑制范式下观察到了人脑额下回区的适应现象。Kilner等的成功也首次为人脑额下回 (inferior frontal gyrus, IFG) (即恒河猴 F5区在人脑中的同源区)中存在镜像神经元提供了有力的证据。
Keysers和 Gazzola (2009a)曾质疑利用单细胞记录技术从癫痫病患者身上获得镜像神经元存在证据的可行性。但 Mukamel等(2010)利用单细胞记录技术,在 21位癫痫病患者的杏仁核、海马、内嗅皮层、海马旁回、辅助运动区、前辅助运动区、前扣带回背侧和边缘位置均发现了具有镜像属性的神经元,并进一步根据它们在各皮层区域的数量分布采取了相应的统计方法,从而确定了各皮层区域中所观察到的这些神经元的分布不是随机的。统计结果表明,海马、海马旁回、内嗅皮层和辅助运动区内具有镜像属性的神经元分布呈显著性水平。这意味着,从统计学的角度看,Mukamel等第一次在单一细胞水平上直接证实了人脑中存在具有镜像属性的神经元。
尽管 Welberg (2010)不认可 Mukamel等的研究成果,认为他们在镜像神经元争议皮层区域以外植入电极,且发现了不属于经典类型的镜像神经元, 这样的结果将会进一步激化关于镜像神经元是否存在的争论。但根据镜像神经元的操作性定义, 以及大量的包括神经成像、经颅磁刺激以及行为研究在内的间接证据,研究者可以大胆推测人脑中确实存在镜像神经元。这种推测还得到来自神经系统种系发生学 (phylogeny)的支持。现有的实验证据显示,鸣禽的高级发声中枢 (HVC)与视觉联合区 (auditory association region, Field L)中存在一类具有镜像属性的神经元 (Prather, Peters, Nowicki, & Mooney, 2008; Keller & Hahnloser, 2009; Roberts, Gobes, Murugan, Olveczky, & Mooney, 2012)。其中 HVC区与猴脑 F5区以及人脑 Broca区具有进化上的同源性。鸣禽 (a)、恒河猴 (b)与人类脑(c)中 “假定的镜像神经元系统” (putative Mirror Neuron System, pMNS)如图 1所示。
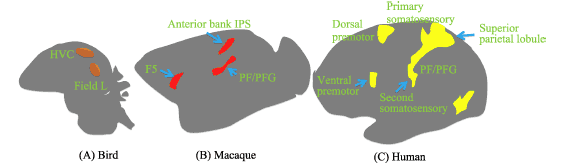
图 1 鸣禽、恒河猴与人类脑中“假定的镜像神经元系统”之比较(改编自:Keysers & Gazzola, 2009b, p. 2)
总之, 当前所有关于人类镜像神经元的研究都是在人脑存在镜像神经元这样一个前提下进行的, 大多数研究结果也倾向于间接证明人脑中镜像神经元的存在,且没有研究结果直接证明人脑中不存在镜像神经元。因此,我们可以认为之前研究的成果构成了证明人脑中存在镜像神经元的收敛性证据。那么,人脑中的镜像神经元又是如何起源的呢?
2.2人脑中镜像神经元的起源
在早期关于镜像神经元活动的模仿理论解释 (simulation-theory interpretation)的争论中,研究者们就已经开始非常关注镜像神经元活动是先天遗传还是后天习得的问题 (Gallese & Goldman, 1998)。尤其是在人们清晰地认识到镜像神经元在社会认知活动中的重要意义时,研究者们探究其背后生成机制的愿望就显得愈发强烈。从最初一些思辩性的假设到现在的实证研究,关于镜像神经元起源问题的争议也开始逐渐集中到适应假设(adaptation hypothesis)和联结序列学习假设 (associative sequence learning, ASL)二者身上。这些争议激发研究者们开始系统考察镜像神经元及其系统的形成。
来自进化立场的观点认为,在种系系统发生史和个体发生学 (ontogenetic)中, 诸如动作再认和观察学习这类对生存至关重要的能力在某种程度上可以成为一种先天的能力 (Borenstein & Ruppin, 2005; Lepage & Théoret, 2007)。Sober (2008)认为, 如果有机体的某一特征有助于其自身实现某种特定的功能,那么这种特征就是一种适应。 Heyes (2010a, 2010b)则根据这一定义将进化立场的观点进一步概括为适应。具体地讲,人脑中的镜像神经元被认为是个体在社会认知活动过程中对动作理解的一种适应,而这种适应的本质就是迫于环境压力下的一种自然选择。根据这种观点,人脑中的镜像神经元可以在神经元水平上表现出遗传倾向。换言之,镜像神经元是生来就有的,而感觉经验和运动经验在镜像神经元的形成过程中只起到辅助促进作用。
然而, 适应性假设却对新动作学习过程中所生成的镜像神经元难以做出有效解释。例如, Cook (2012)的研究发现工具使用及视听镜像神经元 (audiovisual mirror neurons)对工具动作和工具声音的反应比对手所执行的抓握以及撕拉景象的反应更大,这种镜像神经元系统对非自然的刺激的更大反应与适应假设是不相符的。但这与 Keysers和 Perrett (2004)、Keysers和 Gazzola (2009a)以及 Giudice, Manera和 Keysers (2009)提出的赫布学习假设(Hebbian learning)相一致。即,镜像神经元是通过限定渠道化的赫布学习 (canalized Hebbian learning)生成的, 这一假设强调基因、环境和经验之间的交互作用在镜像神经元发展过程中的重要性。同时,它并不否认镜像神经元的发展存在遗传倾向,超越了先天和后天之争,并且与现在的进化生物学研究结论相一致。从这一方面而言,镜像神经元的属性可能类似于新热带区域的鸟嘴形态学特征, 它们不仅因觅食和食物处理 (非社会功能, 类似于视觉运动能力 )的需要而被选择,同时也会对歌声的产生 (社会功能, 类似于动作理解 )有所影响(Derryberry et al., 2012)。
赫布学习假设发展至今已逐渐成为联结序列学习假设的一部分。二者最大的区别是前者主要是在微观神经元水平进行探讨,后者则是在宏观水平直接将外显的模仿行为和镜像神经元的活动联系起来。联结学习普遍存在于脊椎动物和非脊椎动物之中,说明这是一种进化上古老且高度保守的对事件之间预测性关系追踪的适应 (Heyes, 2012)。联结序列学习假设认为,个体在发展过程中, 通过将相同动作的观察经验和执行经验建立起某种联系以形成感觉运动经验。这种感觉运动经验在镜像神经元的发展中起着主导性的作用,镜像神经元正是通过重复观察和执行相类似动作的感觉运动经验来生成。镜像神经元更多是一种联结学习过程的副产品,它并不用于实现某种特定功能,而是可以在社会认知活动的不同适应功能中发挥作用(Heyes, 2001, 2010a; Heyes, Bird, Johnson, & Haggard, 2005; Hill et al., 2013)。
继承了进化观点的适应假设和延续了赫布学习的联结序列学习假设也不代表先天和后天之争, 两者都承认遗传和经验对镜像神经元发展的促进作用, 但侧重点不同。适应假设对镜像神经元定义中的特征进行了解释,即镜像神经元是通过遗传进化而获得对观察行为和类似执行行为进行匹配的能力。相比之下,联结序列学习假设则更强调感觉运动经验在镜像神经元发展过程中所起的关键性作用。
研究者们试图通过对新生儿 (猴)的动作模仿的研究来检验上述争论 (Ferrari et al., 2006; Lepage & Théoret, 2007; Jones, 2009; Ray & Heyes, 2011)。由于新生儿很少有机会获得感觉运动经验,因此, 来自新生儿镜像神经元系统的证据能够有力地支持适应性假设。然而,也有一些来自新生儿或婴儿的研究是支持联结学习假设的。 McEwen等(2007)对两岁双胞胎的研究表明模仿方面的个体差异主要源于环境而非遗传。近来的一项研究也发现 1个月时联结学习能力的个体差异能够预测 8个月之后的模仿表现 (Reeb-Sutherland, Levitt, & Fox, 2012)。目前,支持联结序列学习假设的证据主要来自基于人类被试且明确发现感觉运动经验能够提高、抵消甚至是反转镜像激活的一些研究。Catmur, Walsh和 Heyes (2007, 2009), Umiltà等(2008), Catmur等(2008)以及 Cavallo, Heyes, Becchio, Bird和 Catmur (2013)假定, 如果镜像神经元的发展依赖于感觉运动学习,那么我们就可以通过感觉运动学习训练来改变已经非常成熟的镜像神经元系统,甚至赋予它们逆向镜像 (counter-mirror)属性。他们在研究中通过对被试进行不匹配的感觉运动学习训练成功地验证了他们的假设。换言之,他们的研究结果表明镜像神经元并非与生俱来的,也不是一经获得就无法改变的, 其发展主要依赖于感觉运动学习。
客观地讲, 由于这些研究中用到了已经成熟的镜像神经元系统,因此并不能确切地证实镜像神经元完全是通过感觉运动学习来获得其自身匹配属性的,但至少可以说明感觉运动经验能够塑造镜像神经元。并且,适应假设在缓冲期这个问题上无法对这些研究结果做出合理解释,主要体现在塑造镜像神经元所需要的时间和镜像神经元变化的程度上 (Giudice et al., 2009; Heyes, 2010a)。在这些研究中,镜像神经元在较短的实验时间里发生了变化却没有出现阻止变化的标志。在 Marshall, Boquet, Shipely和 Young (2009)的研究中, 他们更是发现对一个平均时间为 5.5秒的观察学习视频以及随后立即进行的一次模仿便可以产生镜像效应。镜像神经元在如此短的时间里得以塑造与适应假设所期待的变化前的缓冲期假设不符。其次, 这些研究结果表明,原有镜像神经元系统中感觉运动经验能够完全转换而非部分改变。Catmur, Mars, Rushworth和 Heyes, (2011)等在最近的一项双脉冲 TMS (paired-pulse TMS)研究中, 通过对被试进行不匹配感觉运动训练使被试原本建立好的镜像激活联结转换成了一种逆向镜像激活联结。同时,他们还进一步证实了逆向镜像效应和原镜像效应中所涉及的脑区相同,再次强有力地证实了镜像神经元源于联结序列学习。
适应假设和联结序列学习假设各自的侧重点不尽相同,似乎也都可以用来解释镜像神经元的起源, 只是当前的一些研究结果在立场上更支持后者(Catmur et al., 2008; Giudice et al., 2009; Hickok & Hauser, 2010; Heyes, 2010a, 2010b; Catmur, 2013; Cooper, Cook, Dickinson, & Heyes, 2013; Cook, Bird, Catmur, Press, & Heyes, 2014)。除了当前得到了更多实证研究成果的支持外,联结序列学习假设也更符合当前具身认知 (embodied cognition)的认识论立场 (陈巍, 郭本禹, 2014)。其次, 镜像神经元可能不像适应性假设所预设的那样起着关键的、专门化的作用,而是在除动作理解外的动作模仿、共情、言语进化等社会认知活动中起着重要作用。最后,相比于适应假设现在还主要停留在思辨性的或者经验性的研究水平上, 联结序列学习假设已将研究视角从一个较宏观的理论预设落实到一个较微观的研究领域,使得镜像神经元相关研究变得易于操作。此外,联结序列学习假设本身也易于验证。这也是许多研究者倾向于支持它的主要原因。然而,必须指出的是,虽然目前的研究证据倾向于支持联结假设,但这些证据本身在逻辑上并没有排他性的直接否定适应假设,后者并没有在支持前者的实验证据中被 “证伪”。换言之,认为镜像神经元的属性可以通过后天感觉运动的联结学习来获得或改变,并不能由此排斥它在种系发生学与个体发生学上作为进化适应性产物的可能性。因此,这二者的争议至今仍然没有定论,它们依旧是当前镜像神经元研究中的焦点问题 (Iacoboni, 2009a; Heyes, 2010b; Oztop, Kawatob, & Arbib, 2013; Cook et al., 2014)。
3关于镜像“外延”的争议
镜像“内涵”的争议是当前镜像神经元研究中的核心内容,它极大地影响着我们对镜像神经元的正确理解。相比之下,有关镜像 “外延”的争议则略显次之。此处的镜像 “外延”主要是指镜像神经元研究的衍生研究,本文主要探讨由镜像神经元系统和镜像机制二者引发的争议。对前者引发的争议进行整理将帮助我们温故镜像神经元研究的发展历程,而对后者进行讨论不单是因为它有望在多数研究中取代 “镜像神经元系统 ”成为描述镜像现象的规范性术语之一,更多的是因为它有望成为短时期内镜像神经元研究的纲领性目标。
3.1人脑中的镜像神经元系统
人脑镜像神经元系统引发的争议源自于 Gallese等(1996)在研究过程中的一个假想。Gallese等根据恒河猴 F5区镜像神经元分类的结果,推断这些镜像神经元将组成一个系统用以匹配所观察到的动作和所执行的动作。在讨论这一系统在动作识别中的作用时,他们假定恒河猴 F5区在人脑中的同源体,即 Brocca区也存在着类似的匹配系统。
Rizzolatti, Fadiga, Matelli等(1996)最早利用 PET技术来精确定位人脑中这种匹配系统所涉及的脑区,以对这一假设进行验证性研究。结果表明, 对抓取动作的观察确实能显著激活人脑皮层中的颞中回和邻近的颞上沟,以及左侧额下回等脑区。随着研究的逐渐展开,有关人脑镜像神经元系统所涉及的相关脑区及镜像网络的争议也迅速蔓延开来。
Iacoboni等 (1999)证实了左侧额叶岛部 (Brodmann 44区)和右侧顶上小叶 (right superior parietal lobule, rSPL)边缘大部在人类动作模仿过程中构成了匹配系统的解剖学基础,这一结果与 Maeda, Kleiner-Fisman和 Pascual-Leone (2002)随后的研究结果一致。 Hamzei等(2003)利用 fMRI技术研究动作再认系统与 Brocca区的关系时发现,无论是在观察还是执行相似行为的过程中,额下回、中央前回、顶叶皮层等脑区均有激活。 Buccino, Binkofski和 Riggio (2004), Baumgaertner, Buccino, Lange, McNmara和 Binkofski (2007)和 de Zubicaray, Postle, McMahon, Meredith和 Ashton (2010)也发现, 人类被试的 Brodmann 44区在其观察和执行手部或者口部相类似动作时也都被激活。
正如上述脑成像研究所表明的,人类在观察与执行相类似的动作时,左侧额下回、颞中回、颞上沟、右侧顶上小叶喙部、中央前回等脑区都有被激活的现象。换言之,人脑的这些区域都有可能是人类镜像神经元系统的重要组成部分。但鉴于大部分研究中都有左侧额下回后部(posterior IFG)和和顶下小叶喙部 (rostral IPL)被激活的证据,Rizzolatti和 Craighero (2004)认为至少这两个区域构成了镜像神经元系统的核心。 Iacoboni和 Dapretto (2006)则进一步主张, 镜像神经元系统涉及到一个核心的环路,它包括三个脑区。其中有两个脑区是镜像神经元网络的组成部分,即额下回后部与腹侧前运动区的连结处和顶下小叶前部。第三个区域是在镜像神经元系统核心区域定义之外的颞上沟后部。这一主张得到多数研究者的支持 (Keysers & Gazzola, 2009a; Rizzolatti & Sinigaglia, 2010; Sinigaglia, 2010; Fogassi & Ferrari, 2010)。
但事实上, 并非所有的研究者都认可左侧额下回后部和顶下小叶喙部在镜像神经元系统中的重要作用。 Grèzes, Armony, Rowe和 Passingham (2003)认为, 前中央沟腹侧边缘可能是恒河猴 F5区对应的人脑中同源体的组成部分,而作为连结前运动皮层和前额叶皮层的过渡性区域,Brodmann 44区只不过构成了人脑 F5区的前端。 Jonas等 (2007)在利用事件相关 fMRI探究对不及物手指动作的观察和空间一致地模仿会引起额下回后部和顶下小叶前部的变化特征时发现,单独观察一个手指运动并不能显著激活这两个区域。他们推测出现这种现象的原因可能是人类顶 -额镜像网络 (parieto-frontal mirror network)对简单不及物动作的响应严重依赖于实验情境。
Molenberghs等(2009)对 20篇探讨这两个区域在动作模仿中的作用的文章进行了基于激活可能性的估计 (activation likelihood estimation, ALE)和基于标签 (label-based)的元分析 (meta-analysis)研究。结果表明,动作模仿中主要涉及的脑区是顶上小叶、顶下小叶和背侧前运动皮层,而非额下回。并且,动作模仿过程中持续激活的是额叶和前运动皮层,也非额下回。换言之,动作模仿中所涉及的脑区,并非经典镜像神经元系统所涉及的脑区。随后,他们再次用相同的方法对 125篇 fMRI研究进行元分析以确定人脑中具有镜像属性的脑区时发现,除上额下回、背侧 /腹侧前运动区皮层、顶上 /下小叶外,镜像属性脑区的精确定位取决于任务形式 (Molenberghs et al., 2012)。另外, Turella, Pierno, Tubaldi和 Castiello (2009)也从理论基础、研究方法、实验设计、结果分析等角度对先前采用 fMRI和 PET技术来研究人脑镜像神经元系统的大量文章进行了严格的再审视。他们认为,当前假定动作观察、动作模仿和动作执行共用了重叠的网络是合理的,但是仅通过比较几个不同的实验结论就简单地得出某些脑区 (如 Brodmann 44区)是镜像神经元系统区域的结论是不合理的。最近,Oosterhof, Tipper和 Downing (2013)采用重复抑制范式以及多维模式分析方法 (multivariate pattern analysis, MVPA)发现了前顶叶区域对于动作知觉的重要作用。此外,他们的研究还进一步为枕颞叶两侧皮质在人类镜像神经元系统中的作用提供了更多的证据。然而,他们的发现却无法为前运动皮层在镜像神经元系统中的作用提供更多的支持。
此外, 来自 EEG/MEG的研究发现,“μ波抑制 (mu rhythm suppression)”现象可以成为检验人类大脑中镜像神经元系统活动的重要生物学指标。即, 动作观察会引起观察者感觉运动皮层内部 μ波节律的抑制性变化,并且这种变化与个体在动作执行过程中出现的 μ波节律变化非常相似,只是变化幅度较小 (Muthukumaraswamy & Johnson, 2004)。进一步来自成人、婴儿与新生猴的 EEG研究发现, 在动作观察过程中的 μ波抑制并非是由于观察者外显的运动系统的变化造成的 (Muthukumaraswamy, Johnson, & McNair, 2004; Nystr.m, Ljunghammar, Rosander和 Von Hofsten, 2010; Ferrari et al., 2012), 且观察非生物性的运动则不会引发 μ波抑制现象 (Oberman, Hubbard, McCleery, Altschuler, Ramachandran, & Pineda, 2005; Oberman, Pineda, & Ramachandran, 2007; Ferrari et al., 2012)。这就意味着 μ波节律的变化是纯粹表征水平而不是激活水平上的大脑皮层活动所导致的。当然,近期也有 EEG-fMRI的同步研究发现,并非所有的镜像神经元系统的区域都严格对应存在动作执行与观察动作中共变的 μ波抑制现象, 例如 Brodmann 44区(Arnstein, Cui, Keysers, Maurits, & Gazzola, 2011; Braadbaart, Williams, & Waiter, 2013)。因此,将μ波抑制视为支持镜像神经元系统活动的直接与可靠证据仍然存在争议 (Vanderwert, Fox, & Ferrari, 2013)。
至此, 我们认为有必要区分在镜像外延研究中经常用到的两个术语:镜像神经元系统(mirror neuron system)和镜像属性区域 (regions with mirroring property)。镜像神经元系统又可分为猴镜像神经元系统与人类镜像神经元系统。猴镜像神经元系统主要由顶下小叶的喙部 (rostral part of the inferior parietal lobule, IPL,即 PF/PFG区)和腹侧前运动皮层 (ventral premotor cortex, vPM, 即 F5区)组成。人类镜像神经元系统则专指具备类似视觉–运动对应关系的运动皮层区域(Rizzolatti & Fabbri-Destro, 2008; Rizzolatti & Sinigaglia, 2010)。结合来自进化神经科学与重复抑制范式 fMRI的证据, 目前普遍认可的人类经典镜像神经元系统只包含两个脑区:额下回的后部和顶下小叶喙部(Rizzolatti & Craighero, 2004; Kilner & Lemon, 2013; Cook et al., 2014)。相比之下,镜像属性区域则泛指一切在自我 .他人的认知活动之间构成直接“镜像匹配 ” (mirror matching)的脑区, 这些区域本身不具备运动性属性,其功能也就大大地拓展到了动作理解之外的诸多社会认知现象(Muthukumaraswamy & Johnson, 2007; Hickok, 2009)。例如, “共情” (empathy)指的是在观察到他人某种情绪时自身也会产生类似的情感体验。作为一个情绪与感觉的加工过程,共情牵扯到诸多的感觉皮层和皮层下结构,不仅仅是感觉运动皮层的功能 (Frith & Frith, 2012)。因此,负责共情的脑区应仅被标以镜像属性区域,而不应直接列入人类镜像神经元系统的组成部分。
综上, 目前看来, 任何对镜像神经元系统所涉及的脑区和网络进行精确描述的尝试都可能是不合时宜的,因而对镜像神经元系统的争议做出评判意义并不大。时下研究者更应该关注的是人脑中的镜像机制,对镜像神经元系统的描述也应该建立在已经弄清镜像机制的基础之上。
3.2人脑中的镜像机制
之所以对当前镜像神经元系统引发的争议不做评判, 一个很重要的原因是因为许多研究对 “镜像神经元系统 ”这个术语本身的谬用。 Heyes (2010a)强调我们在使用镜像神经元系统这一术语时要谨慎,只不过她主要是从镜像神经元起源的角度来考虑。镜像神经元系统是动作模仿、动作再认和动作理解等复杂认知活动的基础。从这种意义上讲,“镜像神经元系统 ”可以被视为一个典型的开放的复杂巨系统 (钱学森,于景元, 戴汝为,1990; 佘振苏,2012), 它包含了不同亚型的镜像神经元系统。之所以称其为 “复杂”的, 是因为这些亚型的镜像神经元系统在功能上既可以特异性地服务于某类社会认知功能(Fabbri-Destro & Rizzolatti, 2008), 也可以表现出有别于经典镜像神经元系统的其它镜像属性 (Mukamel et al., 2010)。之所以称其为是“开放”的, 是因为就目前的研究而言,对镜像神经元系统的功能与镜像属性的了解正处于证据积累阶段。未来完全可能发现新的镜像神经元系统亚型或子系统。相反,那些将镜像神经元系统笼统地等同于镜像属性区域的研究本身就值得商榷。
就单个亚型的镜像神经元系统而言,它不仅包括各类镜像神经元,还包括复杂的镜像机制 (Buccino et al., 2004; Iacoboni et al., 2005; Oztop et al., 2013), 即一种统一动作知觉和动作执行的机制, 能削弱个体观察和自身卷入状态之间的差异 (Rizzolatti & Sinigaglia, 2010; Sinigaglia, 2010)。因而, 对镜像机制的研究和直接验证某些脑区是否为镜像神经元系统组成部分的研究还是有着本质的区别。 Pacherie和 Dokic (2006)提到要区分镜像神经元系统和镜像机制,但他们更关注镜像神经元的属性和功能,所以只区分了广义和狭义的镜像神经元系统,并未对镜像神经元的工作机制做出进一步解释。
按照钱学森对开放的复杂巨系统从定性到定量的研究思路,时下镜像神经元研究在短期内应以镜像机制研究为主。这样既可以使我们的研究变得可行,又可以使研究对象更加明确,同时还可以为我们进一步对镜像神经元系统描述工作的展开打下良好基础。
最早明确提出将镜像机制作为研究对象的是 Iacoboni等(1999), 他们证实在人脑皮层中存在一种将观察到的动作与这个动作的内部运动表征进行直接匹配 (direct-matching)的机制,而动作模仿正是基于这种直接匹配机制。 Rizzolatti, Fogassi和 Gallese (2001)随后也再次证实这种镜像机制的存在。但 Buccino等(2001)认为, 这种匹配机制匹配的是所观察动作的感觉表征和动作表征库中的运动表征。当 Iacoboni等(1999)等在研究中仅要求被试去观察动作而不进行模仿时,并未发现顶上小叶有明显激活,且在动作观察过程中左侧额下回的激活是由将观察动作内部词语化 (internal verbalization)所造成的。因此,这种直接匹配机制并非镜像机制。镜像神经元是否促进动作理解,仍然存在较大的争议(Hickok, 2013)。
Rizzolatti和 Craighero (2004)发现除镜像机制外, 可能还有其它神经机制在动作理解和动作再认中发挥重要作用。 Fabbri-Destro和 Rizzolatti (2008)再次对镜像机制在动作和意图理解中的作用做了回顾,结果发现,镜像机制跟随镜像神经元在人脑皮层中位置的变化而变化,它们分别在意图理解、模仿,语言和情绪体验等社会认知活动中发挥重要作用。同时,他们还强调人脑中可能存在许多不同的镜像神经元系统,当前已有证据初步指向上述设想 (Rizzolatti & Fogassi, 2014)。
Iacoboni和 Dapretto (2006)在人脑额叶皮层的镜像神经元中观察到了一种脉冲率变化的现象, 有超过 80%的镜像神经元在动作执行中表现出兴奋性反应而在动作观察中则表现出抑制性反应,剩下小部分镜像神经元则表现出了完全相反的形式。他们称这种小部分表现出相反兴奋和抑制形式的镜像神经元为 “超级镜像神经元 ” (super mirror neurons)。超级镜像神经元被认为处于功能性神经元层,是经典镜像神经元的最高层,因而能够控制和调节较低水平镜像神经元的活动 (Iacoboni, 2008, 2009b)。这种超级镜像神经元在区别自生成表征状态和通过动作观察所生成的表征状态中起着重要作用,可以帮助我们在动作执行中保持主体感并且抑制动作观察中不必要的模仿(Mukamel, Ekstrom, Kaplan, Iacoboni, & Fried, 2007; Mukamel et al., 2010)。简言之,这个两个镜像神经子系统可以帮助我们灵活地整合和区分自己和他人动作的知觉和运动方面。进一步的,Wang, Ramsey和 Hamilton (2011)的 fMRI研究发现, 在模仿行为发生过程中,镜像神经元系统的活动受到内侧前额皮层 (medial prefrontal cortex, mPFC)的实时调控。当环境中包含的社会性信号需要个体进行人际交流时,内侧前额皮层就会促进镜像神经元系统的活动,加强无意识模仿 ; 而当环境中出现交流停止的社会性信号时,内侧前额皮层就会抑制镜像神经元系统的活动,从而阻断对观察动作的无意识模仿。因此,在这种情况下, 镜像神经元系统又成为了上述 “社会性自上而下的反应调控机制” (social top-down response modulation, STORM) (Wang & Hamilton, 2012)的子系统。
综上所述, 我们当前对镜像机制的认识还非常模糊。尤其是在 Iacoboni和 Mukamel等发现了不同于传统经典镜像机制的另一种镜像机制,Wang等进一步澄清外部情境对其的作用之后,对镜像机制的具体特征进行深入研究就显得更为迫切。如果要对镜像神经元系统做进一步的研究,就必须弄清各种类型的镜像机制及其相互关系。
4 镜像神经元研究展望
一个非常重要的事实是,自 di Pellegrino等首次发现镜像神经元的这 20余年来,有关镜像神经元的研究已广泛开展开来,其中不乏出色的研究成果,但也存在着诸多争议。镜像神经元研究本身的复杂性以及间接性,使得常用的单细胞记录、PET、TMS、fMRI和 ERP等技术显得并不完美。因此, 在科学研究的认识论层面上,必须警惕将镜像神经元视为 “认知科学的圣杯 ” (Holy Grail of Cognitive Science)这样一种极端偷懒的神经还原主义倾向。在方法论层面上,对于相关的实验设计和结论解释需要小心谨慎。尤其是,在尽可能明晰镜像神经元操作性定义的前提下,相关实验设计的内部效度必须被尽快提上日程。
虽然, 当前镜像神经元的研究具有上述不可避免的不利因素,但并不影响其继续深入下去。整合现有的镜像神经元研究已有成果,我们认为,镜像神经元将来的研究重点可能会出现如下趋势:(1)通过改变未来该领域的提问方式以推进对镜像神经元功能与镜像机制的认识。传统研究中类似“镜像神经元是用来做什么?”或“它们是为什么服务的? ”这样的询问方式本身就是存在着过分依赖适应假说的风险。至少研究者还应该同时询问“赋予镜像神经元以 ‘镜像’(mirrorness)过程是怎么样的,或者说镜像神经元所具有的这种可以将观察和执行动作匹配起来的首要功能是如何产生的? ” (Cook et al., 2014)。在这种提问方式下,Kohler等(2002)和 Keysers等(2003)在恒河猴的前运动皮层发现的视听镜像神经元以及 Ferrari, Rozzi和 Fogassi (2005), Rochat等(2010)在豚尾猴 F5区外侧相继发现的各种“工具响应型镜像神经元” (tool-responding mirror neurons)等各类 “逻辑相关的镜像神经元(logically-related mirror neurons), 以及超级镜像机制都有可得到更系统的解释。当然, 这也意味着旧有镜像神经元研究范式要做出相应调整,尤其是采用 fMRI技术的研究。 (2)同时考察各种镜像机制及其与其他社会认知的神经机制之间的关系。未来的研究不仅需要积极探究人脑不同皮层区域在意图理解、动作模仿、语言和情绪体验等社会认知功能中的作用,以及镜像神经元系统与这些皮层区域联合活动实现某种社会认知功能的具体机制 (Keysers, 2009)。例如,如果感觉–运动属性是镜像神经元的诸多功能的首要属性, 那么这种低阶的神经表征机制如何与非感觉运动 (nonsensorimotor)的、抽象的高阶神经表征之间产生联系,从而实现各种社会认知活动(Caramazza et al., 2014)。又如, 理解非意图的动作 (non-intended actions)不仅需要镜像神经元系统的参与, 而且需要在时间与空间范畴内对突发事件的信号进行编码,这就需要右侧颞顶联结 (right temporo-parietal junction, rTPJ), 左侧缘上回 (left supramarginal gyrus) 和内侧前额皮层的共同参与 (Buccino, Baumgaertner, Colle, Buechel, Rizzolatti, & Binkofski, 2007)。(3)在发展科学 (Developmental Science)视角下审视并检验镜像神经元的功能与起源问题。从发展科学的视角来看,上述问题实际上是“天性–教养”之争(The Nature-Nurture Debates)在社会认知活动及其神经基础上的延续与回响。因此, Heyes (2014)建议遵循动物行为学先驱 Niko Tinbergen的学说, 从因果关系 (causation)、个体发生学(ontogeny)、存在价值 (survival value)与进化 (evolution)四个方面来系统考察镜像神经元的功能与起源问题。同样的, 在这方面, 郭任远和 Gottlieb的渐成论 (Epigenesis)思想也具有重要启示作用。 (4)以一些特殊群体作为研究对象检验镜像神经元研究中的争议。比如,利用婴儿来研究镜像神经元的个体发生学特征 ; 利用神经外科患者来进行深度植入的单细胞记录研究 ; 利用永久植物状态或最小意识状态的个体来探究经典镜像神经元系统、镜像属性区域与其它社会脑 (social brain)区域之间的关系 (Chen, Shan, Guo, & Yuan, 2013); 利用考察联觉者 (synaesthetes)对他人躯体感觉的体验来检测躯体感觉皮层与镜像神经元系统之间的复杂联结关系 (Keysers, Kaas, & Gazzola, 2010)等, 都可以帮助克服我们当前研究中一些不可回避的缺陷。当然,这必须在道德、法律和患者本人知情同意的前提下展开。
总之, 即便近年来的研究已经逐步将镜像神经元请下了 “科学神坛 ”, 也并不妨碍其成为改变认知神经科学的重要发现 (Brass & Rüschemeyer, 2010)。当然,由于镜像神经元结构与功能的复杂性, 决定了其所带来的争议必将持续下去。最后,正如 Cook等(2014)建议的那样: “关于镜像神经元功能的可靠信息,只能基于发展的历史、系统的理论以及谨慎的实验而获得,同时我们也需要理智地去审视这些争议。”
参考文献
陈巍, 郭本禹 . (2014). 具身.生成的认知科学 : 走出 “战国时代”.心理学探新, 34(2), 111–116.
钱学森, 于景元, 戴汝为. (1990).一个科学新领域——开放的复杂巨系统及其方法论. 自然杂志, 13(1), 3–10. 佘振苏. (2012). 复杂系统学新框架 ——融合量子与道的知识体系. 北京: 科学出版社. Arnstein, D., Cui, F., Keysers, C., Maurits, N. M., & Gazzola,
V. (2011). μ-suppression during action observation and execution correlates with BOLD in dorsal premotor, inferior parietal, and SI cortices. The Journal of Neuroscience, 31(40), 14243–14249.
Baumgaertner, A., Buccino, G., Lange, R., McNmara, A., & Binkofski, F. (2007). Polymodal conceptual processing of human biological actions in the left inferior frontal lobe. European Journal of Neuroscience, 25(3), 881–889.
Borenstein, E., & Ruppin, E. (2005). The evolution of imitation and mirror neurons in adaptive agents. Cognitive Systems Research, 6, 229–242.
Borg, E. (2007). If mirror neurons are the answer, what was the question? Journal of Consciousness Studies, 14(8), 5–19.
Borg, E. (2013). More question for mirror neurons. Consciousness and Cognition, 22(3), 1122–1131.
Braadbaart, L., Williams, J. H. G., & Waiter, G. D. (2013). Do mirror neuron areas mediate mu rhythm suppression during imitation and action observation? International Journal of Psychophysiology, 89, 99–105.
Brass, M., & Rüschemeyer, S.-A. (2010). Mirrors in science: How mirror neurons changed cognitive neuroscience. Cortex, 46(1), 139–143.
Brown, E. C., & Brüne, M. (2012). The role of prediction in social neuroscience. Frontiers in Human Neuroscience, 6,
Buccino, G., Baumgaertner, A., Colle, L., Buechel, C., Rizzolatti, G., & Binkofski, F. (2007). The neural basis for understanding non-intended actions. Neuroimage, 36, T119–T127.
Buccino, G., Binkofski, F., & Riggio, L. (2004). The mirror neuron system and action recognition. Brain and Language, 89, 370–376.
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L., Gallese, V.… Freund, H. J. (2001). Action observation activates premotor and parietal areas in a somatotopic manner: An fMRI study. European Journal of Neuroscience, 13, 400–404.
Caggiano, V., Pomper, J. K., Fleischer, F., Fogassi, L., Giese, M., & Their, P. (2013). Mirror neurons in monkey area F5 do not adapt to the observation of repeated actions. Nature Communications, 4, 1433.
Caramazza, A., Anzelotti, S., Strnad, L., & Lingnau, A. (2014). Embodied cognition and mirror neurons: A critical assessment. Annual Review of Neuroscience, 37, 1–15.
Casile, A. (2013). Mirror neurons (and beyond) in the macaque brain: An overview of 20 years of research. Neuroscience Letters, 540, 3–14.
Catmur, C. (2013). Sensorimotor learning and the ontogeny of the mirror neuron system. Neuroscience Letters, 540, 21– 27.
Catmur, C., Gillmeister, H., Bird, G., Liepelt, R., Brass, M., & Heyes, C. (2008). Through the looking glass: Counter- mirror activation following incompatible sensorimotor learning. European Journal of Neuroscience, 28, 1208– 1215.
Catmur, C., Mars, R., Rushworth, M., & Heyes, C (2011). Making mirrors: Premotor cortex stimulation enhances mirror and counter-mirror motor facilitation effects. Journal of Cognitive Neuroscience, 23(9), 2352–2362.
Catmur, C., Walsh, V., & Heyes, C. (2007). Sensorimotor learning con.gures the human mirror system. Current Biology, 17, 1527–1531.
Catmur, C., Walsh, V., & Heyes, C. (2009). Associative sequence learning: The role of experience in the development of imitation and the mirror system. Philosophical Transactions of the Royal Society B: Biological Sciences, 364, 2369–2380.
Cavallo, A., Heyes, C., Becchio, C., Bird, G., & Catmur, C. (2013). Timecourse of mirror and counter-mirror effects measured with transcranial magnetic stimulation. Social Cognitive and Affective Neuroscience, 9, 1082–1088.
Chen, W., Shan, C. L., Guo, B. Y, & Yuan, T. F. (2013). Mirror neuron system activation for conscious states assessment: For patients in vegetative and minimally conscious states. Medical Hypotheses, 81, 144–145.
Chong, T. T. J., Cunnington, R., Williams, M. A., Kanwisher, N., & Mattingley, J. B. (2008). fMRI adaptation reveals mirror neurons in human inferior parietal cortex. Current Biology, 18(20), 1576–1580.
Cook, R. (2012). The ontogenetic origins of mirror neurons: Evidence from ‘tool-use’ and ‘audiovisual’ mirror neurons. Biology Letters, 8(5), 856–859.
Cook, R., & Bird, G. (2013). Do mirror neurons really ‘mirror’ and do they really code for action goals? Cortex, 49, 2944–2945.
Cook, R., Bird, G., Catmur, C., Press, C., & Heyes, C. (2014). Mirror neurons: From origin to function. Behavioral and Brain Science, 37(2), 177–241.
Cooper, R. P., Cook, R., Dickinson, A., & Heyes, C. M. (2013). Associative (not Hebbian) learning and the mirror neuron system. Neuroscience Letters, 540, 28–36.
de Zubicaray, G., Postle, N., McMahon, K., Meredith, M., & Ashton, R. (2010). Mirror neurons, the representation of word meaning, and the foot of the third left frontal convolution. Brain and Language, 112(1), 77–84.
Derryberry, E. P., Seddon, N., Claramunt, S., Tobias, J. A., Baker, A., Aleixo, A., & Brumfield, R. T. (2012). Correlated evolution of beak morphology and song in the neotropical woodcreeper radiation. Evolution, 66(9), 2784–2797.
di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G. (1992). Understanding motor events: A neurophysiological study. Experimental Brain Research, 91(1), 176–180.
Dinstein I., Hasson, U., Rubin, N., & Heeger, D. J. (2007). Brain areas selective for both observed and executed movements. Journal of Neurophysiology, 98, 1415–1427.
Dinstein, I., Thomas, C., Behrmann, M., & Heeger, D. J. (2008). A mirror up to nature. Current Biology, 18, R13–R18. Dinstein, I. (2008). Human cortex: Reflections of mirror
neurons. Current Biology, 18(20), 956–959.
Eshuis, R, Coventry, K. R., & Vulchanova, M. (2009). Predictive eye movements are driven by goals, not by the mirror neuron system. Psychological Science, 20(4), 438–440.
Fabbri-Destro, M., & Rizzolatti, G. (2008). Mirror neurons and mirror systems in monkeys and humans. Physiology, 23, 171–179.
Fan, Y. T., Decety, J., Yang, C. Y., Liu, J. L., & Cheng, Y. (2010). Unbroken mirror neurons in autism spectrum disorders. Journal of Child Psychology and Psychiatry, 51, 981–988.
Ferrari, P. F. Vanderwert, R., Paukner, A., Bower, S., Suomi, S., & Fox, N. (2012). Distinct EEG amplitude suppression to facial gestures as evidence for a mirror mechanism in newborn monkeys. Journal of Cognitive Neuroscience, 24(5), 1165–1172.
Ferrari, P. F., Rozzi, S., & Fogassi, L. (2005). Mirror neurons responding to the observation of actions made with tools in monkey ventral premotor cortex. Journal of Cognitive Neuroscience, 17(2), 212–226.
Ferrari, P. F., Visalberghi, E., Paukner, A., Fogassi, L., Ruggiero, A., & Suomi, S. J. (2006). Neonatal imitation in rhesus macaques. PLoS Biology, 4(9), e302.
Fogassi, L., & Ferrari, F. P. (2010). Mirror systems. Wiley Interdisciplinary Reviews: Cognitive Science, 2(1), 22–38.
Frith, C. D., & Frith, U. (2012). Mechanisms of social cognition. Annual Review of Psychology, 63, 287–313.
Gallese, V., & Goldman, A. (1998). Mirror neurons and the simulation theory of mind-reading. Trends in Cognitive Sciences, 2(12), 493–501.
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in the premotor cortex. Brain, 119, 593–609.
Gallese, V., Gernsbacher, M., Heyes, C., Hickok, G., & Iacoboni, M. (2011). Mirror neuron forum. Perspectives on Psychological Science, 6(4), 369–407.
Gazzola, V., Aziz-Zadeh, L., & Keysers, C. (2006). Empathy and the somatotopic auditory mirror system in humans. Current Biology, 16, 1824–1829.
Giudice, M. D., Manera, V., & Keysers, C. (2009). Programmed to learn? The ontogeny of mirror neurons. Developmental Science, 12(2), 350–363.
Glenberg, A. M. (2011). Introduction to the mirror neuron forum. Perspectives on Psychological Science, 6(4), 363–368.
Grèzes, J., Armony, J. L., Rowe, J., & Passingham, R. E. (2003). Activations related to “mirror” and “canonical” neurons in the human brain: An fMRI study. Neuroimage, 18, 928–937.
Grill-Spector, K., & Malach, R. (2001). fMR-adaptation: A tool for studying the functional properties of human cortical neurons. Acta Psychologica, 107, 293–321.
Hamilton, A. F. de C. (2013). Reflecting on the mirror neuron system in autism: A systematic review of current theories. Developmental Cognitive Neuroscience, 3, 91–105.
Hamilton, A. F. de C., & Grafton, S. T. (2006). Goal representation in human anterior intraparietal sulcus. The Journal of Neuroscience, 26(4), 1133–1137.
Hamzei, F., Rijntjes, M., Dettmers, C., Glauche, V., Weiller, C., & Buchel, C. (2003). The human action recognition system and its relationship to Broca’s area: An fMRI study. Neuroimage, 19(3), 637–644.
Heyes, C. (2001). Causes and consequences of imitation. Trends in Cognitive Science, 5(6), 253–261.
Heyes, C. (2010a). Where do mirror neurons come from. Neuroscience and Biobehavioral Reviews, 34, 575–583.
Heyes, C. (2010b). Mesmerising mirror neurons. NeuroImage, 51, 789–791.
Heyes, C. (2012). Simple minds: A qualified defence of associative learning. Philosophical Transactions of the Royal Society B: Biological Sciences, 367(1603), 2695–2703.
Heyes, C. M. (2014). Tinbergen on mirror neurons. Philosophical Transactions of the Royal Society B: Biological Sciences, 369, doi: 10.1098/rstb.2013.0180
Heyes, C., Bird, G., Johnson, H., & Haggard, P. (2005). Experience modulates automatic imitation. Cognitive Brain Research, 22, 233–240.
Hickok, G. (2009). Eight problems for the mirror neuron theory of action understanding in monkeys and humans. Journal of Cognitive Neuroscience, 21(7), 1229–1243.
Hickok, G. (2013). Do mirror neurons subserve action understanding? Neuroscience Letters, 540, 56–58.
Hickok, G., & Hauser, M. (2010). (Mis)understanding mirror neurons. Current Biology, 20(14), 593–594.
Hill, A. T., Fitzgibbon, B. M., Arnold, S. L., Rinehart, N. J., Fitzgerald, P. B., & Enticott, P. G. (2013). Modulation of putative mirror neuron activity by both positively and negatively valenced affective stimuli: A TMS study. Behavioural Brain Research, 249, 116–123.
Hutchison, W., Davis, K., Lozano, A., Tasker, R., &
Dostrovxky, J. (1999). Pain related neurons in the human cingulate cortex. Nature Neuroscience, 2, 403–405. Iacoboni M. (2008). Mesial frontal cortex and super mirror neurons. Behavioral and Brain Science, 31(1), 30. Iacoboni, M. (2009a). Imitation, empathy, and mirror neurons. Annual Review of Psychology, 60, 653–670.
Iacoboni M. (2009b). Mirroring people: The new science of how we connect with others (2nd). New York, USA: Farrar, Straus & Giroux.
Iacoboni, M., & Dapretto, M. (2006). The mirror neuron system and the consequences of its dysfunction. Nature Review Neuroscience, 7, 942–951.
Iacoboni, M., Molnar-Szakacs, I., Gallese, V., Buccino, G., Mazziotta, J. C., & Rizzolatti, G. (2005). Grasping the intentions of others with one’s own mirror neuron system. PLoS Biology, 3(3), e79.
Iacoboni, M., Woods, P. R., Brass, M., Bekkering, H., Mzaaiotta, J. C., & Rizzolatti, G. (1999). Cortical mechanisms of human imitation. Science, 286, 2526–2528.
Jonas, M., Siebner, H. R., Biermann-Ruben, K., Kessler, K., B.umer, T., Büchel, T. … Münchau, A. (2007). Do simple intransitive finger movements consistently activate frontoparietal mirror neuron areas in humans. Neuroimage, 36, T44–T453.
Jones, S. S. (2009). The development of imitation in infancy. Philosophical Transactions of the Royal Society B: Biological Sciences, 364, 2325–2335.
Keller, G. B., & Hahnloser, R. H. (2009). Neural processing of auditory feedback during vocal practice in a songbird. Nature, 457, 187–190.
Keysers, C. (2009). Mirror neurons: Where can I find out more?. Current Biology, 19(21), R971–R973.
Keysers, C., & Gazzola, V. (2009a). Unifying social cognition. In J. A. Pineda (Ed.), Mirror neuron systems: The role of mirroring processing in social cognition (pp. 3–39). New York, USA: Human Press.
Keysers, C., & Gazzola, V. (2009b). Expanding the mirror: Vicarious activity for actions, emotions and sensations. Current Opinions in Neurobiology, 19, 1–6.
Keysers, C., & Gazzola, V. (2010). Social neuroscience: Mirror neurons recorded in humans. Current Biology, 20(8), R353–R354.
Keysers, C., & Perrett, D. I. (2004). Demystifying social cognition: A Hebbian perspective. Trends in Cognitive Science, 8(11), 501–507.
Keysers, C., H. Kaas. J., & Gazzola, V. (2010). Somatosensation in social perception. Nature Reviews Neuroscience, 11, 417–428.
Keysers, C., Kohler, E., Umiltà, M. A., Nanetti, L., Fogassi, L., & Gallese, V. (2003). Audiovisual mirror neurons and action recognition. Experimental Brain Research, 153, 628–636.
Kilner, J. M., & Lemon, R. N. (2013). What we know currently about mirror neurons. Current Biology, 23(23), R1057–R1062.
Kilner, J. M., Neal, A., Weiskopf, N., Friston, K. J., & Frith,
C. D. (2009). Evidence of mirror neurons in human inferior frontal gyrus. The Journal of Neuroscience, 29(32), 10153–10159.
Kohler, E., Keysers, C., Umiltà, M. A., Fogassi, L., Gallese, V., & Rizzolatti, G. (2002). Hearing sounds, understanding actions: Action representation in mirror neurons. Science, 297, 846–848.
Kosonogov, V. (2012). Why the mirror neurons cannot support action understanding. Neurophysiology, 44(6), 499–502.
Lepage, J. F., & Théoret, H. (2007). The mirror neuron system: Grasping others actions from birth. Developmental Science, 10(5), 513–529.
Lingnau, A., Gesierich, B., & Caramazza, A. (2009). Asymmetric fMRI adaptation reveals no evidence for mirror neurons in humans. Proceedings of the National Academy of Science of the United States of America, 106(24), 9925–9930.
Maeda, F., Kleiner-Fisman, G., & Pascual-Leone, A. (2002). Motor facilitation while observing hand actions: Speci.city of the effect and role of observer’s orientation. Journal of Neurophysiology, 87, 1329–1335.
Marshall, P. J., Boquet, C. A., Shipely, T. F., & Young, T. (2009). Effects of brief imitative experience on EEG desynchronization during action observation. Neuropsychologia, 47(10), 2100–2106.
McEwen, F., Happe, F., Bolton, P., Rijsdijk, F., Ronald, A., Dworzynski, K., & Plomin, R. (2007). Origins of individual differences in imitation: Links with language, pretend play, and socially insightful behavior in two-year- old twins. Child Development, 78(2), 474–492.
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2009). Is the mirror neuron system involved in imitation? A short review and meta-analysis. Neuroscience and Biobehavioral Reviews, 33, 975–980.
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2012). Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies. Neuroscience and Biobehavioral Reviews, 36, 341–349.
Mukamel, R., Ekstrom, A. D., Kaplan, J., Iacoboni, M., & Fried, I. (2007). Mirror properties of single cells in human medial frontal cortex. In the Annual Meeting of the Society for Neuroscience (37th) (Abstract, 127, 4). San Diego.
Mukamel, R., Ekstrom, A. D., Kaplan, J., Iacoboni, M., & Fried, I. (2010). Single-neuron responses in humans during execution and observation of actions. Current Biology, 20(8), 750–756.
Muthukumaraswamy, S. D., & Johnson, B. W. (2004).
Changes in rolandic mu rhythmduring observation of a precision grip. Psychophysiology, 41, 152–156.
Muthukumaraswamy, S. D., & Johnson, B. W. (2007). A dual mechanism neural framework for social understanding. Philosophical Psychology, 20, 43–63.
Muthukumaraswamy, S. D., Johnson, B. W., & McNair, N.
A. (2004). Mu rhythm modulation during observation of an object-directed grasp. Cognitive Brain Research, 19, 195–201.
Nystr.m, P., Ljunghammar, T., Rosander, K., & Von Hofsten, C. (2010). Using mu-rhythm perturbations to measure mirror neuron activity in infants. Developmental Science, 14, 327–335.
Oberman, L. M., Hubbard, E. M., McCleery, J. P., Altschuler,
E. L., Ramachandran, V. S., & Pineda, J. A. (2005). EEG evidence for mirror neuron dysfunction in autism spectrum disorders. Cognitive Brain Research, 24, 190–198.
Oberman, L. M., Pineda, J. A., & Ramachandran, V. S. (2007). The human mirror neuron system: A link between action observation and social skills. Social Cognitive and Affective Neuroscience, 2(1), 62–66.
Oosterhof, N. N., Tipper, S. P., & Downing, P. E. (2013). Crossmodal and action-specific: Neuroimaging the human mirror neuron system. Trends in Cognitive Sciences, 17(7), 311–318.
Oztop, E., Kawatob, M., & Arbib, M. A. (2013). Mirror neurons: Functions, mechanisms and models. Neuroscience Letters, 540, 43–55.
Pacherie, E., & Dokic, J. (2006). From mirror neurons to joint actions. Cognitive Systems Research, 7, 101–112.
Prather, J, S., Peters, S., Nowicki, S., & Mooney, R. (2008). Precise auditory-vocal mirroring in neurons for learned vocal communication. Nature, 451, 305–310.
Ramachandran, V. S. (2011). The tell-tale brain: A neuroscientist's quest for what makes us human. New York: W. W. Norton & Company.
Ray, E., & Heyes, C. (2011). Imitation in infancy: The wealth of the stimulus. Developmental Science, 14(1), 92–105.
Reeb-Sutherland, B. C., Levitt, P., & Fox, N. A. (2012). The predictive nature of individual differences in early associative learning and emerging social behavior. PLoS One, 7(1), e30511.
Rizzolatti G., & Craighero, L. (2004). The mirror-neuron system. Annual Review of Neuroscience, 27, 169–192.
Rizzolatti, G., & Luppino, G. (2001). The cortical motor system. Neuron, 31, 889–901.
Rizzolatti, G., & Sinigaglia, C. (2010). The functional role of the parieto-frontal mirror circuit: Interpretations and misinterpretations. Nature Reviews Neuroscience, 11, 264–274.
Rizzolatti, G., Fadiga, L., Gallese, V., & Fogassi, L. (1996). Premotor cortex and the recognition of motor actions. Cognitive Brain Research, 3, 131–141.
Rizzolatti, G., Fadiga, L., Matelli, M., Bettinardi, V., Paulesu, E., Perani, D., & Fazio, F. (1996). Localization of grasp representations in humans by PET: 1. Observation versus execution. Experimental Brain Research, 111, 246–252.
Rizzolatti, G., Fogassi, L., & Gallese, V. (2001). Neurophysiological mechanisms underlying the understanding and imitation of action. Nature Reviews Neuroscience, 2(9), 661–670.
Rizzolatti, G., & Fogassi, L. (2014). The mirror mechanism: Recent findings and perspectives. Philosophical Transactions of the Royal Society B: Biological Sciences, 369, doi: 10.1098/rstb.2013.0420
Roberts, T. F., Gobes, S. M. H., Murugan, M., Olveczky, B. P., & Mooney, R. (2012). Motor circuits are required to encode a sensory model for imitative learning. Nature Neuroscience, 15, 1454–1459.
Rochat, M. J., Caruana, F., Jezzini, A., Escola, L., Intskirveli, I., Grammont, F., … Umiltà, M. A. (2010). Responses of mirror neurons in area F5 to hand and tool grasping observation. Experimental Brain Research, 204, 605–616.
Shmuelof, L., & Zohary, E. (2005). Dissociation between ventral and dorsal fMRI activation during object and action recognition. Neuron, 47, 457–470.
Sinigaglia, C. (2010). Mirroring and making sense of others. Nature Reviews Neuroscience, 11, 449.
Sober, E. (2008). Evidence and evolution: The logic behind the science. New York, USA: Cambridge University Press.
Turella, L., Pierno, C. A., Tubaldi, F., Castiello, U. (2009). Mirror neurons in humans: Consisting or confounding evidence. Brain and Language, 108, 10–21.
Uithol, S., Van Rooij, I., Bekkering, H., & Haselager, W. F. G. (2011). What do mirror neurons mirror? Philosophical Psychology, 24(5), 607–623.
Umiltà, M. A., Escola, L., Intskirveli, I., Grammont, F., Rochat, M., Caruana, F., … Rizzolatti, G. (2008). When pliers become fingers in the monkey motor system. Proceedings of the National Academy of Sciences of the United States of America, 105, 2209–2213.
Vanderwert, R. E., Fox, N. A., & Ferrari, P. F. (2013). The mirror mechanism and mu rhythm in social development. Neuroscience Letters, 540, 15–20.
Wang, Y., & Hamilton, A. F. de. C. (2012). Social top-down response modulation (STORM): A model of the control of mimicry in social interaction. Frontiers in Human Neuroscience, 6, 153.
Wang, Y., Ramsey, R, Hamilton, A. F. de, C. (2011). The control of mimicry by eye contact is mediated by medial prefrontal cortex. The Journal of Neuroscience, 31, 12001–12010.
Welberg, L. (2010). Mirrors, mirrors, everywhere. Nature Reviews Neuroscience, 11, 374–374.
Connotations and Extensions of “Mirrorness”: The Controversies on Mirror Neurons
CHEN Bo1,2; CHEN Wei3,4; ZHANG Jing4; YUAN Tifei5
(1 Scientific Instruments Department, ISEN Tech. & Trading Co., Ltd, Beijing 100086, China) (2 Center for Cognition and Brain Disorders, Hangzhou Normal University, Hangzhou 310015, China) (3 Department of Psychology, Shaoxing University, Shaoxing, 312000, China) (4 Center for Language and Cognition Research, Zhejiang University, Hangzhou, 310028, China)
(5 School of Psychology, Nanjing Normal University, Nanjing 210097, China) Abstract: Mirror neurons are considered to be the important findings which had been changing cognitive neuroscience. They provide a uniform perspective of study for lots of mind or cognitive phenomena which can’t be tested by experiments yet. Their crucial role in social cognition leads to researchers’ extensive attentions and drastic controversies. The authors review the main arguments according to the logic relations and properties, and divide them into two categories: one is about the mirror connotations, including the disputations in whether the human brain contains mirror neurons and their origin; the other one focuses mainly on the mirror extensions consisting of the contentions of mirror neuron system and mirror mechanism. The future investigations must abandon the tendency of reductionism that regards mirror neurons as “Holy Grail of Cognitive Science”, but consider them as open complex giant systems which are consisted of multiple sub systems, and transfer our attentions to the mirror mechanism. Key words: mirror neurons; mirror connotations; mirror extensions; mirror neuron system; mirror mechanism
www.psychspace.com心理学空间网

