
依恋的神经生理机制*
陈文凤 王争艳 王 岩
(首都师范大学教科院心理系,北京 100083)
摘 要 在动物社会依恋的神经生物学研究基础上,研究者对人类依恋的神经生理机制进行了初步探讨,一致的结论是无论婴儿还是成人,不安全依恋个体的心率、皮质醇水平高于安全依恋的个体。而在依恋系统激活时婴儿和成人的脑电活动和大脑的功能定位研究却发现了不一致:较之安全依恋婴儿,不安全依恋婴儿的左侧前额大脑区域显示了较少的活动;成人依恋类型的脑机制研究并无定论。所有这些研究多为相关研究,今后有待于在神经生物水平上对依恋的基本理论问题如内部工作模式的意识和无意识性、母婴依恋与成人依恋的关系(特别是一般依恋表征和特殊依恋表征)以及母婴依恋对婴幼儿大脑发育的影响等问题展开深入研究。
关键词 依恋;自主神经系统;下丘脑-垂体-肾上腺;脑
分类号 B844;B845
对社会依恋神经生理机制的探讨最初来自对哺乳动物母性行为和雌雄交配的研究,特别是在啮齿类动物和灵长类动物依恋的精神生物学研究方面,成果更是丰硕。
在母性行为研究方面,研究者通常以鼠和猴为研究对象,主要关注母亲看护中幼崽自主神经系统和内分泌系统的变化。鼠模型是动物母婴依恋研究的基础,其中最有意义的研究结果是,早期生活中相对单一的紧张性刺激会使内分泌系统发生变化。例如,幼鼠在出生两周内,与母鼠短暂分离(15分钟)会导致血液中皮质类固醇物质(corticosteroid)的升高,这种物质对分离焦虑有缓解作用。若与母鼠的分离超过 3 个小时,幼鼠体内血浆促肾上腺皮质激素(plasma adrenocorticomimetic hormone,ACTH)和肾上腺酮(corticosterone)的浓度将增加,直接的行为反应是幼鼠可能会对未来的刺激反应比较敏感(Putnam, 2005)。
母亲看护对幼体的内分泌、神经生长和大脑发育影响重大。研究人员发现:得到母亲高水平看护的幼鼠其特定受体和海马类胆碱能神经的分布(与学习和记忆有关)明显增加。在灵长类动物模型的研究中,通过对比母猴抚养的猴子和同伴喂养的猴子,Suomi 发现它们在面对短暂分离时血液化学物质、活动方式、肾上腺皮质、神经化学和行为反应方面存在大量差异(Putnam, 2005)。也就是,母猴喂养的猴子比与同伴一起长大的猴子对环境具有更强的适应性。
在动物交配研究方面,研究者发现,催产素对雌雄依恋有促进作用。由垂体分泌的催产素不仅对乳汁分泌和子宫收缩有重要的作用,更能促进雌雄交配。Carter在依恋形成的生物模型中指出,与依恋对象的身体接触或维持亲近可以引发催产素释放,而催产素可降低下丘脑-垂体-肾上腺(hypothalamic -pituitary-adrenal,HPA)反应和交感神经系统(sympathetic nervous system,SNS)的反应,增加副交感神经系统(parasympathetic nervous system,PNS)的反应(Carter, 1998),从而降低个体的焦虑。
与动物研究相似,依恋理论认为婴儿面对现实可能的威胁和危险时会向照看者寻求亲近以获得安全感(Bowlby, 1982)。在婴儿与母亲的分离阶段,作为婴儿安全港湾的母亲一旦离开,对婴儿来说是一种危险的体验,在婴儿感知到外界环境对自身的威胁时,会自动激活依恋系统(Bowlby, 1982),与此同时也会使神经内分泌系统和大脑处于激活状态。由于安全型、回避型和矛盾型婴儿的依恋行为模式存在很大差异,危险引发的神经生理反应也应该存在差异。早期依恋研究者最初测量人类母婴依恋时就很注重对不同依恋类型的生理反应差异的研究。后来,Hazan和Shaver(1987)把依恋研究拓展到成人,即婚恋依恋,在恋人之间也存在相互依赖的情感联结,伴侣也给另一方提供安全的港湾。在成人研究中,依恋焦虑和依恋回避个体在压力条件下有不同的反应,依恋焦虑的个体为了维持依恋系统的激活容易产生和夸大消极情绪状态;回避的个体容易忽视危险事件,抑制可能激活依恋系统的与危险相关的想法或情绪。那么不同依恋类型个体的神经机制有怎样的差异?这些差异又是如何测得的呢?本文将对依恋的神经生理机制及其相关的研究做一总结与回顾。
1 依恋与自主神经系统
1.1 理论基础
自主性神经系统(autonomic nervous system,ANS)是表达压力反应的主要系统,它的功能是支配那些不受意识控制的组织和器官,如心肌。自主性神经系统包括交感神经系统和副交感神经系统。下丘脑对自主神经系统具有调节作用。下丘脑后侧区具有使血压升高,心率加快等交感神经反应的功能;下丘脑前侧区则具有使血压下降,心率减慢等副交感神经反应的功能。所以心率、血压可以很灵敏地反映自主神经反应,而且交感神经系统和副交感神经系统具有相互拮抗的作用(Diamond, 2001)。
危险和压力在激活自主神经系统的同时也激活了依恋系统,而依恋系统的核心功能是安全基地和缓解痛苦(Bowlby, 1982)。Shaver等人认为与依恋对象的安全的依恋经验和成功地缓冲压力,对个体有效情绪调节策略的发展有很大影响(Fox & Card, 1999)。因而依恋关系对个体在压力条件下的情感和生理反应有调节作用。三者之间的具体关系可以用图1来表示。
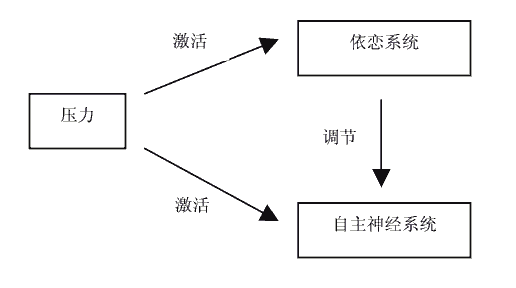 图1 依恋与自主神经系统的关系示意图
图1 依恋与自主神经系统的关系示意图
1.2 相关研究
心率和血压都可以反映自主神经系统的活动,但鉴于心率测量的可操作性和简洁性,在依恋生理测量中多用心率作为主要生理指标。许多研究测查了陌生情境中分离和重聚阶段不同依恋类型婴儿的心率变化情况,但是结论并不一致。Fox和Card(1999)在综述中提到,有的研究发现,安全型、回避型和矛盾型婴儿在分离和重聚阶段的心率都有所升高;而另外的研究表明,在陌生情境中安全型和不安全型依恋的婴儿在分离阶段有着相同的心率,而在重聚阶段又有所不同。最近的研究则发现,回避型的儿童在分离阶段尽管没有表现出过度痛苦的信号,但心跳是加速的,并且在重聚后没有像以往研究那样发现依恋类型之间的差异(Zelenko, Kraemer, Huffman, Gschwendt, Pageler, & Steiner, 2005)。可见,这方面的结果和结论很不统一,需要进行进一步的研究。
相比婴儿,成人压力状态下心率的测量取得了较为一致的研究结果。Carpenter 和 Kirkpatrick(1996)考察了大学女生在两种不同的实验室压力情境下的生理反应,一种是恋人在场,一种是恋人不在场,不同的依恋类型表现出了不同的生理反应:当她们恋人不在场的时候,回避型的个体显示了更高的血压和心率活动。这表明个体对依恋对象可得性的期待调节着压力的情绪反应;而安全依恋的个体不管恋人是否在场,没有表现出对压力的不同的反应,说明安全依恋的个体可以将来自依恋对象的压力缓冲功能进行成功地内化,所以就不会依靠依恋对象在场的具体的支持。也就是说,是依恋对象在个体生活中的出现而不仅仅是在实验室中的出现缓冲了个体的压力。
心率、血压等生理指标参数多是由交感和副交感神经双重支配的,而心率的加快,可能是交感神经活动增强引起的,也可能是副交感神经减弱的结果(阎克乐, 张文彩, 张月娟, 封文波, 袁立壮, 王兰爽 等,2006)。如果要单纯反映交感神经或副交感神经的活动,心率变异性的频谱分析是比较理想的指标。心率变异性(heart rate variability,HRV)指的是心跳快慢的变化情况,由于心跳快慢是由两个相邻的R-R间期时间(心室一次兴奋到下一次兴奋所用的时间)长短决定的,因此,研究者们由R-R间期的变化计算出一系列参数,规定的两个主要的参数有低频成分(LF);高频成分(HF)。高频(HF)是由迷走神经单独介导;低频(LF)受迷走神经和交感神经共同调制,但主要反映交感神经的活动性(阎克乐 等,2006)。
Robert, William, Robert, Jonathan 和 David(2006)用心率变异性方法研究了成人的不安全依恋与主观压力和诱发压力的关系。研究通过让成人自由回忆压力事件和实验者设计的压力任务引发被试情绪后测量5个阶段(基线、压力事件回忆阶段、放松、压力任务、放松)被试的心率。通过心率变异性频谱分析发现,在压力事件回忆阶段,依恋焦虑与压力事件有显著的正相关;在压力任务阶段,依恋回避分数在高频(HF)HRV和低频(LF)HRV上有显著的主效应。说明依恋焦虑与主观的压力测量相联系,而依恋回避与压力任务相联系。
心率的变异性受到迷走神经的调控,而迷走神经中主要成份之一为副交感神经。副交感神经的退缩比活动更能促进心率活动。而迷走神经中的起伏现象可以在压力条件下观察到。Diamond 和 Angela(2005)的研究发现,迷走神经的活性与依恋焦虑呈负相关,与依恋安全性呈正相关。在依恋关系中感受到更多安全的人在经历了实验室诱发的愤怒后,恢复更快,心跳和呼吸更快回到静态水平。但并没有证实婚恋依恋对象的出现对迷走神经在情绪恢复中的调节中起作用。最近的一项研究考察了母亲的生理调节是否可以调节婴儿负性情绪与母亲敏感性的关系。研究发现对于回避型婴儿的母亲,迷走神经退缩与母亲对婴儿痛苦的敏感性有关;而对安全依恋婴儿的母亲,没有发现迷走神经退缩与母亲敏感性的关系(Mills-Koonce, Cox, & Propper, 2007)。
2 依恋与神经-内分泌系统
2.1 理论基础
内分泌系统是机体重要的调节系统,它与神经系统一起调节机体活动,维持体内平衡。目前在依恋研究领域,研究者关注最多的是下丘脑-垂体-肾上腺(HPA)轴对人类依恋系统的调节作用。对动物和人类的研究发现,HPA对社会刺激十分敏感,HPA的反应和调节受早期社会经历的影响(Gunnar & Donzella, 2002)。而HPA反应大多发生在危险的环境中,而这个危险的环境也会激活依恋系统,所以它就成了研究依恋很好的指标(Wang, Bartolome, & Schanberg, 1996)。HPA轴对压力的反应可参看具体过程如下:外部压力刺激作用于下丘脑,增加促皮质激素释放激素(corticotropin releasing hormone, CRH)的分泌,HPA轴被激活,CRH通过循环从下丘脑到垂体前叶,它刺激促肾上腺皮质激素(adrenocorticotropic hormone, ACTH)的分泌。循环的ACTH刺激肾上腺皮质分泌肾上腺皮质激素,对于人类主要是分泌皮质醇。皮质醇是肾上腺在应激反应里产生的一种类激素,可以平衡身体的压力反应。肾上腺血压的增加可以反馈给下丘脑受体和垂体腺体,从而降低CRH和ACTH分泌,以达到降低压力反应的作用。
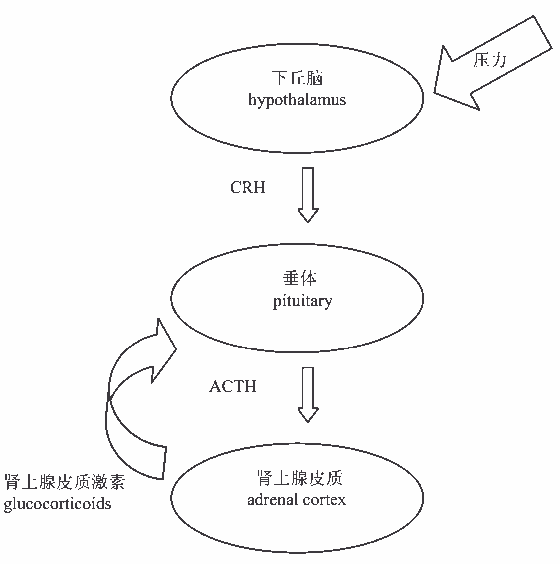 图2 HPA轴对压力的反应图示(Putnam, 2005)
图2 HPA轴对压力的反应图示(Putnam, 2005)
注:ACTH表示促肾上腺皮质激素,CRH表示促皮质素释放激素
对鼠和猴的研究都发现,与母亲的接触可以降低HPA的压力反应。Hennessy, Hornschuh, Kaiser和Sachser(2006)对土拨鼠生命全程的研究验证了皮质醇的社会缓冲作用。压力状态下身体需要皮质醇来维持正常的生理机能,如果没有皮质醇,身体将无法对压力作出有效反应。下面我们将具体看一下这方面的相关研究。
2.2 相关研究
在进行依恋研究时,经常通过对皮质醇水平进行分析,探讨皮质醇对压力的调节作用。皮质醇的收集可以通过唾液、血浆或尿液获得,但考虑到实验的无创伤性和方便性,进行依恋研究时一般采取收集唾液的方式。由于唾液中皮质醇相当稳定不易失活,因此,在实验中收集到的唾液可以精确反映下丘脑-垂体-肾上腺(HPA)反应。在家庭背景下和在陌生情境中收集唾液的时间有所差异。在家庭中一般是在婴儿起床后和睡觉前收集;由于皮质醇需要一段反应时间,因而在陌生情境后的 15~30分钟内收集到的唾液可以反映HPA反应。
皮质醇对压力的调节和缓冲作用得到了来自婴幼儿依恋类型和照看质量研究的支持。陌生情境中婴儿各种依恋类型差异的研究发现,与父母不安全依恋关系婴儿的皮质醇水平比与父母有安全依恋关系的婴儿的皮质醇水平要高(Gunnar, 2002; Schieche & Spangler, 2000)。这表明在陌生情境中安全型依恋婴儿比不安全型依恋婴儿能更好地处理分离带来的痛苦。对学步儿的研究也发现,不安全依恋的儿童比安全依恋的儿童在受到惊吓和痛苦时更可能显示出皮质醇的升高(Spangler & Schieche, 1998)。
父母或看护人的敏感性和支持性以及母亲的抑郁情绪、父母婚姻质量等家庭因素都会影响儿童的皮质醇水平。Pendry 和 Adam(2007)在自然生态背景下探讨了父母婚姻功能、母亲照料质量和母亲的情绪功能与儿童皮质醇水平的关系。结果表明低质量的婚姻满意度、频繁的言语攻击与儿童高的皮质醇水平相关。但临时照看者也可以在一定程度上作为父母的补充,对儿童的压力情绪起到缓解作用。Gunnar等人发现敏感的、反应型的临时照看者同样可以缓解儿童与母亲分离导致的皮质醇的升高。
对领养儿童的研究发现,受到更多忽视的儿童在早上有较低的皮质醇水平,并且皮质醇水平从早到晚很少会有变化(Dozier, Manni, Gordon, Peloso, & Gunnar, 2006)。而正常儿童的皮质醇代谢遵循则这样一种生理节奏:一般皮质醇水平最高在早晨(约6~8点),最低点在凌晨(约0~2点)。通常在上午 8点~12 点间皮质醇水平会骤然下跌,之后全天都持续一个缓慢的下降趋势。Fisher, Stoolmiller 和Gunnar(2007)通过对被领养儿童12个月的干预研究发现,早期教养质量的提高对预防HPA轴功能失调具有显著的影响。
总之,看护者的照看质量影响着儿童皮质醇的变化,从而直接影响到儿童对压力的反应和调节能力,进而影响儿童的身心发展。
另外,Gunnar(2002)指出,在亲密关系中,皮质醇水平对于社会压力和支持更为敏感。Filipp等考察了男性和女性在没有社会支持、有陌生人的社会支持和有伴侣的社会支持条件下的皮质醇活动,发现,对男性而言,伴侣的支持与皮质醇的下降有很大关系,与依恋对象作为压力调节者的结论一致;然而,对女性而言,伴侣支持与更高的皮质醇活动相联系,得出了与以前研究不一致的结果(Kirschbaum, Klauer, Filipp, 1995)。这就对皮质醇的压力缓冲作用提出了质疑。而 Powers, Pietromonaco, Gunlicks和Sayer(2006)探讨了恋人的依恋类型与人际冲突时皮质醇反应的关系,发现不安全依恋的个体比安全依恋的个体在面对人际冲突(实验室任务)时显示了高水平的压力反应;拥有不安全伴侣的男性比起拥有安全伴侣的男性在面对人际冲突时也显示了高水平的压力反应,而没有发现拥有不安全和安全伴侣的女性皮质醇水平的差异。看来性别与依恋类型的交互作用是今后值得探讨的一个话题。
3 依恋与脑
3.1 理论基础
依恋是母亲与婴儿之间的一种情感联结,母亲看护是良好依恋关系建立的关键。母亲看护对婴儿的情绪和心理的影响已被广泛证实,而对婴儿的大脑发育的影响越来越引起研究者的关注。Bowlby(1982)在描述依恋系统的社会和生物功能时阐述了右半球的作用。因为右半球与边缘系统和皮层下区域存在着广泛的联系,所以它在情绪信息加工和调节上起主导作用。目前的神经心理研究也显示,婴儿的情绪经历在大脑发展阶段不成比例地存储并且在右半球加工,婴儿右半球的成熟受早期社会经验的影响比较显著。依恋关系形成的内部工作模式经过加工存储在右脑皮层的内隐记忆系统中。特别是与母亲面对面情感交流时,母亲面部表情的输入可以诱发婴儿大脑某种营养因子的产生,例如脑衍生神经营养因子(brain-derived neurotrophic factor,BDNF)。母亲看护可以增加兴奋性氨基酸(N-methyl-D-aspartate,NMDA)受体水平,提高脑衍生神经营养因子水平和增加婴儿大脑突触。BDNF是促进中脑多巴胺神经的因子,而可以激活NMDA受体的多巴胺在大脑皮层的发展中起着促进作用(Schore, 2001)。
母亲对婴儿的大脑发育有如此重要的影响,那么母亲在与婴儿的互动中大脑又会有怎样的变化呢?许多文章都论述了母亲与婴儿的依恋经验和面对面情感交流直接影响了眶额皮层的电回路。眶额皮层是边缘系统所属成分之一。Bowlby认为,边缘系统与依恋行为的唤醒和发展变化有关。边缘系统主要负责对面部表情进行无意识的加工,并可以迅速作出调整以适应环境。另外,眶额皮层具有无意识倾向,能够在意识发生之前对未来行为的重要程度进行编码,并对信息加工和行为选择机制相联系的位置进行表征。眶额皮层与积极情感的评价有关,在母婴依恋中起着重要作用(Nitschke, Nelson, Rusch, Fox, Oakes, & Davidson, 2004)。
3.2 相关研究
随着神经科学的日趋发展和高科技手段的日益完善,关于依恋的脑机制的研究取得了丰硕的成果,许多研究用脑电图(electroencephalograph,EEG)、事件相关电位(Event-related Potentials,ERP)、功能磁共振成像( functional magnetic resonance imaging,fMRI)等技术对依恋的脑机制进行了探讨,下面我们将分别介绍婴儿和成人在这些方面的相关研究。
研究者最初的兴趣是婴儿在与母亲互动中的大脑半球偏侧化问题。大量的研究表明,左额叶的激活与接近行为和表达积极情感的能力有关。右额叶的激活与回避行为和表达消极情感的能力有关。右额叶表现出激活增强的婴儿更可能在分离时表现出悲伤,并很难控制悲伤;左额叶表现出激活增强的婴儿更可能表现出积极情感,并能更好的控制行为反应(Fox, 1999)。Dawson, Ashman, David和Susan(2001)用脑电仪记录了婴儿在陌生情境中的活动。与安全型依恋的婴儿相比,不安全依恋婴儿的左侧前额大脑区域显示了较少的活动。这反映出其采取了退缩的情绪调节策略。
许多研究用fMRI考察了母亲对婴儿(自己的孩子和别人的孩子)的声音(哭和笑)和图片(婴儿照片)刺激反应的大脑活动,发现了不同脑区的激活(Leibenluft, Gobbini, Harrison, & Haxby, 2004; Ranote, Elliott, Mtchell, Deakin, & Appleby, 2004)。例如,Noriuchi, Kikuchi和Senoo(2008)用fMRI考察了母亲在观看婴儿微笑和哭两种条件的录像片段(没有声音)时母亲的大脑反应,结果表明:在辨别自己的孩子时,母亲的眶额叶皮质、中脑导水管、前脑岛、背侧和腹外侧核区域被激活;而在看到婴儿哭泣时则表现出了不同的激活区域:眶额皮层的背侧区域、尾壳核、右侧额下回、前额皮质、前扣带回、后扣带回、丘脑、颞上沟。
Shaver等人也用fMRI检测了不同依恋风格成人在思考情绪事件和企图抑制这些想法时大脑的激活模型,发现:依恋焦虑与情绪相关的脑区(如颞极)呈正相关,与情绪调节的区域(眶额皮质)呈负相关,这表明焦虑型的人在回忆失去经历时不能调节消极情绪;回避型的人在两个脑区(扣带回皮层,侧面前额皮层)显示了更多的去活化(Shaver, 2005)。这一结论可以用Shaver就依恋风格与情绪调节策略的关系提出的依恋综合控制模型来解释:依恋焦虑的人采用过度激活策略,这种策略以一种慢性激活状态维持着依恋系统,可能导致个体对危险事件的夸大从而集中注意于危险事件。所以,焦虑依恋的个体为了维持依恋系统的激活容易产生和夸大消极情绪状态。回避型的个体以一种不活跃的状态来维持依恋系统,即去活化策略,容易忽视危险事件,并抑制可能激活依恋系统的与危险相关的想法或情绪(Mikulincer & Shaver, 2003)。
Goldstein等人用ERP技术研究了依恋风格如何调整自己的大脑反应以适应不同的情绪刺激(Zilber, Goldstein, & Mikulincer, 2007)。结果表明那些在依恋焦虑维度上得分较高的被试比在依恋焦虑维度上得分低的被试在观看消极图片时引发更多的晚期正向电位。先前的研究报告了依恋相关的情绪对行为,认知的影响,并把他们解释为是由过度激活或去活化策略引起的,但这些策略的描述仅限于抽象或理论层面,对他们的认知和脑机制的研究还很少。而ERP对依恋定向的测量可以建立高水平的理论构建和低水平的认知机制的桥梁。
4 前景展望
依恋概念自提出以来就一直受到发展心理学家、临床心理学家以及人格/社会心理学家的关注,有关神经生理机制的探讨也从未间断过。随着生物学、医学及神经科学的发展,对依恋心理机制和生理机制的探讨也日益深入。生理测量以生物学为依托,借助精确的测量,探讨依恋的生理机制,该方面研究的开展无论是对依恋的问卷测量研究还是访谈研究都是非常有益的补充,它可以在一定程度上搭建发展心理学研究趋向和人格/社会心理学研究趋向相互沟通的桥梁,改善长期以来两种研究趋向各自为阵的局面,对依恋的基本理论问题如内部工作模式的意识和无意识性、母婴依恋与成人依恋的关系、成人的一般依恋表征和特殊依恋表征以及母婴依恋对婴儿大脑发育的影响等问题做出更为深入的解答。
在生物学文献中有一个一般假设,依恋的机制对以下所有方面都是一致的:婴儿对母亲,母亲对婴儿,成年女性对成年男性以及成年男性对成年女性,社会依恋是生物适应性的一种。情爱和母爱两者都有稳固的情感联系,有着类似的进化论根源,有着相似的生物学功能,很可能有共同的神经机制(Insel & Young, 2001)。Curtis和Wang(2003)也认为母婴依恋和成人依恋有同样的神经激素基础,只是存在位置和性别的特定性。那么母婴依恋和成人依恋是否真有类似的脑机制?对母婴依恋和成人依恋共同生理机制的探讨是一个有价值的方向。目前的许多研究多是分别探讨母婴依恋或者成人依恋的脑功能定位,只有极少的研究同时探讨了两者的脑功能定位问题。如Bartels和Zeki(2004)用fMRI测了母亲在观看婴儿和恋人图片时的大脑活动,发现两种依恋激活的脑区在壳核、苍白球和尾状体三个部位有重叠。但是,其他纵向研究表明,成人依恋表征并不能由婴儿期的依恋类型得到预测(Grossmann, 2005),所以Curtis的假设是否正确,还值得相关的纵向研究的探讨。
依恋的脑电研究取得了一定的成果,深化了我们对依恋的脑机制的理解,为我们今后的研究提供了新的视角,但这些研究多停留在描述的、相关研究阶段,对于依恋的脑功能定位还没有一致的结论。在更深的神经化学水平上探讨早期母婴依恋对婴幼儿脑发育的影响,这种早期情感依恋在脑内的印刻对以后情绪管理(特别是极端的焦虑、抑郁等)的作用,探寻成人后脑内的早期情感依恋的印刻物(不同的神经递质的水平变化及相关感受器的分布密度)以及它的神经回路等的研究将是非常有意义的。今后,借助人类脑成像的各种技术手段的结合使用,如通过使用弥散张量成像(Diffusion tensor imaging,DTI)非侵入性的跟踪技术和功能磁共振成像,记录不同脑区间白质的联结的同时给以灰质定位,可能会比较清楚的描述出情感依恋的神经回路。
危险与压力的程度、突发性特点会影响神经生理水平,也会影响个体的认知和情感(Diamond, 2001)。面对突如其来的刺激(比如尖叫)个体可能来不及做出认知上的评价就会产生强烈的生理反应;而对可预期的刺激(比如陌生人出现)个体可以对其作出认知评价,这在一定程度上可以缓解生理反应。依恋系统本身也具有压力缓解的作用。所以个体的认知和情感都对生理起调节作用。认知因素和情感因素对生理的调节是否存在程度上的差异或者三者之间存在中介效应?对于此的探讨会使依恋的研究更加深入。
此外,在临床研究方面,可以考虑把依恋相关的生理指标作为评估依恋干预研究效果的指标。但是目前的研究多是相关研究,生理测量都只是作为情绪唤醒的客观指标,并没有探索与依恋联系的基本生理特性的更深层次的问题。混乱型依恋日益受到儿童心理学家的关注,因为如果这种依恋类型儿童发展不好将导致病理性依恋,从而影响其人格的发展。所以探讨混乱型依恋和病理性依恋的生理机制和脑机制有利于对其成因的分析,从而为其干预和治疗提供依据。
参考文献
内分泌系统 . 来自互联网 : http://baike.baidu.com/view
/92172.htm
阎克乐, 张文彩, 张月娟, 封文波, 袁立壮, 王兰爽 等.
(2006). 心率变异性在心身疾病和情绪障碍研究中的应用.
心理科学进展, 14(2), 261–265.
Bartels, A. & Zeki, S. (2004). The neural correlates of maternal
and romantic love. NeuroImage, 21(3), 1155–1166.
Bowlby, J. (1982). Attachment and loss: Attachment. New York:
Basic Books.
Carpenter, E. M., & Kirkpatrick, L. A. (1996). Attachment style
and presence of a romantic partner as moderators of
psychophysiological responses to a stressful laboratory
situation. Personal Relationships, 3(4), 351–367.
Carter, C. S. (1998). Neuroendocrine perspective on social
attachment and love. Psychoneuroendocrinology, 23(8),
779–818.
Curtis, J. T., & Wang, Z. X. (2003). The neurochemistry of pair
bonding. Current Directions in Psychological Science, 12(2),
49–53.
Dawson, G., Ashman, S. B., David, H., & Susan, S. (2001).
Autonomic and brain electrical activity in securely- and
insecurely-attached infants of depressed mothers. Infant
Behavior and Development, 24(2), 135–149.
Diamond, L. M. (2001). Contributions of Psychophysiology to
Research on Adult Attachment: Review and Recommendations.
Personality and Social Psychology Review, 5(4), 276–295.
Diamond, L. M., & Angela, M. H. (2005). Attachment style,
current relationship security, and negative emotions: The
mediating role of physiological regulation. Journal of Social
and Personal Relationships, 22(4), 499–518.
Dozier, M., Manni, M., Gordon, M. K., Peloso, E., & Gunnar, M.
R. (2006). Foster children’s diurnal production of cortisol: an
exploratory study. Child Maltreatment, 11(2), 189–197
Fisher, P. A., Stoolmiller, M., & Gunnar, M. R. (2007). Effects of
a therapeutic intervention for foster preschoolers on diurnal
cortisol activity. Psychoneuroendocrinology, 32(8-10),
892–905.
Fox, N. A., & Card, J. A. (1999). Psychophysiological measures
in the study of attachment. In Cassidy, J., Phillip, R., & Shaver,
P. R. Handbook of Attachment: Theory, Research, and Clinical
Applications (pp. 226–245). New York: Guilford Press.
Grossmann, K. (2005). Early care and the roots of attachment and
partnership representations: The Bielefeld and Regensburg
longitudinal studies. In Grossmann K E, Grossmann K, Waters
E(Eds.), Attachment from infancy to adulthood(pp. 98–136).
New York: The Guilford Press.
Gunnar, M. R., & Donzella, B. (2002). Social regulation of the 第17 卷第2 期 依恋的神经生理机制 -383-
cortisol levels in early human development. Psychoneuro-
endocrinology, 27(1-2), 199–220.
Hazan, C., & Shaver, P. R.(1987). Romantic love conceptualized
as an attachment process. Journal of Personality and Social
Psychology, 52, 511–524.
Hennessy, M. B., Hornschuh, G., Kaiser, S., & Sachser, N. (2006).
Cortisol responses and social buffering: A study throughout the
life span. Hormones and Behavior, 49(3), 383–390.
Insel, T. R., & Young, L. J. (2001). The neurobiology of
attachment.Nature Reviews Neuroscience, 2 (2), 129–136.
Kirschbaum, C., Klauer, T., Filipp, S. H. (1995). Hellhammer, D.
H. Sex-specific effects of social support on cortisol and
subjective responses to acute psychological stress.
Psychosomatic Medicine, 57(1), 23–31.
Leibenluft, E., Gobbini, M. I., Harrison, T., & Haxby, J. V.
(2004). Mothers’ neural activation in response to pictures of
their children and other children. Biological Psychiatry, 56(4),
225–320.
Mikulincer, M., & Shaver, P. R. (2003). The Attachment
Behavioral System In Adulthood: Activation, Psychodynamics,
And Interpersonal Processes. Advances in Experimental Social
Psychology, 35, 53–152.
Mills-Koonce, W. R., Cox, M., & Propper, C. (2007). Infant and
parent factors associated with early maternal sensitivity: A
caregiver-attachment systems approach. Infant Behavior and
Development, 30(1), 114–126.
Noriuchi, M., Kikuchi, Y., & Senoo, A. (2008). The functional
neuroanatomy of maternal love: mother’s response to infant’s
attachment behaviors. Biological Psychiatry, 63(4), 415–423.
Orbitofrontal cortex tracks positive mood in mothers viewing
pictures of their newborn infants. Neuroimage, 21(2) ,583–692.
Pendry, P., & Adam, E. K. (2007). Associations between parents’
marital functioning, maternal parenting quality, maternal
emotion and child cortisol levels. International Journal of
Behavioral Development, 31(3), 218–231.
Powers, S. I., Pietromonaco, P. R., Gunlicks, M., & Sayer, A.
(2006). Dating couples’ attachment styles and patterns of
cortisol reactivity and recovery in response to a relationship
conflict. Journal of Personality and Social Psychology, 90(4),
613–618.
Putnam, F. W. (2005). The developmental neurobiology of
disrupted attachment: lessons from animal models and child
abuse research. In Berlin, L. J., Amaya-Jackson, L., &
Greenberg, M. T. (Eds.), Enhancing Early Attachments: Theory,
Research, Intervention, and Policy (pp.79–99). New York:
Guilford Press.
Ranote, S., Elliott, R., Mtchell, R., Deakin, J. F. W., & Appleby,
L. (2004). The neural basis of maternal responsiveness to
infants: an fMRI study. Neuroreport, 15(11), 1825–1839.
Robert, G. M., William, J. L., Robert, P. N., Jonathan, J. H., &
David, W. T.(2006). The relationship of attachment insecurity
to subjective stress and autonomic function during standardized
acute stress in healthy adults. Journal of Psychosomatic
Research, 60(3), 283–290.
Schieche, M., & Spangler, G. (2000, July). The moderating role
of maternal sensitivity and attachment security for
biobehavioral organization in infants and toddlers.
Presentation at the International Conference on Infant Studies.
Brighton.
Schore, A. N. (2001). Effects of a secure attachment relationship
on right brain development, affect regulation and infant mental
health. Infant Mental Health Journal, 22(1-2), 7–66.
Shaver, P. R. (2005). Attachment-style differences in the ability
to suppress negative thoughts: Exploring the neural correlates.
NeuroImage, 28(4), 835–847.
Spangler, G., & Schieche, M. (1998). Emotional and
adrenocortical responses of infants to the strange situation: The
differential function of emotional expression. International
Journal of Behavioral Development, 22(4), 681–706.
Wang, S., Bartolome, J. V., & Schanberg, S. M. (1996). Neonatal
deprivation of maternal touch may suppress ornithine
decarboxylase via downregulation of the proto-oncogenes
c-myc and max. The Journal of Neuroscience, 16, 836–842.
Zilber, A., Goldstein, A., & Mikulincer, M. (2007). Adult
attachment orientations and the processing of emotional
pictures-ERP correlates. Personality and Individual Differences,
43(7), 1898–1907.
Zelenko, M., Kraemer, H., Huffman, L., Gschwendt, M., Pageler,
N., & Steiner, H. (2005). Heart rate correlates of attachment
status in young mothers and their infants. Journal of the
American Academy of Child and Adolescent Psychiatry, 44(5),
470–601.
Neurophysiological Mechanism of Human Attachment
CHEN Wen-Feng; WANG Zheng-Yan; WANG Yan
(Department of Psychology, Capital Normal University, Beijing 100048, China)
Abstract: Attachment researchers investigated the neurophysiological mechanism of human attachment based on
neurobiology studies of animal social attachment. They made a conclusion that the insecure individuals have higher
heart rate and high cortisol comparing with the secure ones , While the location of brain function and
electrophysiological study shows that compared to secure infants, insecure infants exhibited less left frontal brain
activities when attachment system was inactivated, but there are no consistent results in adults. However, the
researches are mostly correlational ones and can not provide causal inferences, moreover, the further researches
about basic physiological characteristic of attachment need to be reinforced and discovered.
Key words: attachment; autonomic nervous system; hypothalamic-pituitary-Adrenal; brain
收稿日期:2008-8-15
*
北京市教委人文社科基金项目和北京市重点实验室(学习与
认知)及北京市重点学科(发展与教育)资助。
通讯作者:王争艳,E-mail:hwzy@263.net

