
【摘要】自尊是个体对自己总体的情感性评价。自尊有助于促进个体的心理健康 , 帮助个体应对威胁 , 具有重要的适应价值。近年来随着社会认知神经科学的兴起 , 关于自尊的认知神经机制的研究日益增多 , 主要涉及三个方面:1)自尊与大脑的结构 (如海马体积、前扣带回等区域的灰质体积 )和功能(静息态下的默认网络活动以及脑区之间的功能联结性 )存在一定的关联 ; 2)自尊调节大脑对威胁的反应 , 与高自尊个体相比 , 低自尊个体面对威胁时产生更强的防御性反应 , 自尊通过影响个体面对威胁时的大脑活动帮助个体更好地应对威胁 ; 3)自我评价过程涉及大脑前额叶、眶额叶、扣带回等多个脑区 , 自尊调节个体在自我评价过程中的大脑活动。总之, 自尊作为个体重要的人格特质 , 与大脑的结构、功能以及活动状态等都具有一定的关系 ; 这些发现在一定程度上揭示了自尊的神经机制, 加深了对自尊及其功能的理解。
【关键词】自尊; 自我评价; 威胁; 内侧前额叶; 前扣带回
【分类号】B845; B848
自尊作为个体对自己总体的情感性评价 (Baumeister, 1998; Brown, 2014), 是心理学领域研究最多的课题之一 (Tesser, 2000)。大量行为研究表明, 自尊具有重要的适应价值:高自尊可以增强个体的自我调适能力 , 帮助个体应对威胁和挫折, 促进个体的心理健康和幸福感等 (Baumeister, Campbell, Krueger, & Vohs, 2003; Cai, Wu, & Brown, 2009; Greenberg et al., 1992; Orth, Robins, & Widaman, 2012; Sowislo & Orth, 2013)。为了加深对自尊的理解, 近年来 , 随着认知神经科学和相关技术 (如事件相关电位 (event-related potential, ERP)、功能性磁共振成像 (functional magnetic resonance imaging, fMRI))的发展, 越来越多的研究借助认知神经科学技术探讨自尊的认知神经机制。本文拟对日益增多的涉及自尊的认知神经研究进行梳理和归纳。我们期望在厘清现有研究脉络的基础上 , 为未来的相关研究提供基础和方向性的指导。本文将从三个方面对现有文献予以整理:自尊是否和大脑的某些结构和功能存在关联 ; 不同自尊水平的个体应对威胁时的大脑反应是否存在差异 ; 不同自尊水平的个体在自我评价过程中的大脑活动是否存在差异。鉴于本文所综述的研究均采用罗森伯格自尊量表(Rosenberg, 1965)对自尊进行测量, 并且多数研究基于此对个体的自尊水平进行了区分, 因此下文中将不再对自尊的测量进行介绍。
1自尊与大脑结构和功能的关联
1.1自尊与大脑结构的关联
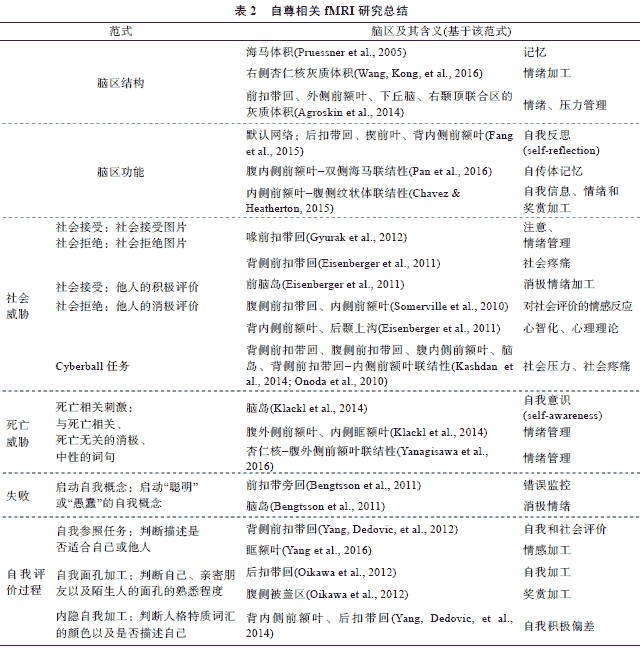
海马(hippocampus)和杏仁核(amygdala)是与自尊相关的重要脑区。海马和杏仁核是大脑边缘系统的重要组成部分 , 分别在记忆和情绪加工中具有重要作用。 Pruessner等(2005)对不同年龄阶段的个体进行大脑扫描 , 发现无论年轻还是年老 , 个体的自尊水平都与其海马体积的大小呈正相关:低自尊个体的海马体积小于高自尊个体。研究者认为这种相关可能有两方面的原因:首先 , 低自尊个体不能合适地对待生活中的消极事件(如社会拒绝 ),对负面信息会产生更强的应激反应(如分泌更多的皮质醇 (cortisol)), 这种持续的应激反应可能导致海马体积萎缩 ; 另一方面 , 由于海马与消极生活事件的记忆情景化有关 , 低自尊个体的海马体积小 , 他们难以正确提取消极事件的情境信息 , 从而容易将消极事件与自我相联系 ; 此外, 在应激情境中 , 他们也难以提取相关的积极记忆(如社会支持 ), 从而导致他们进一步形成消极的自我评价 (杨娟, 张庆林, 2010)。新近的一项研究则发现自尊与海马、杏仁核的结构都存在相关。该研究表明 , 罗森伯格自尊量表的总分与海马灰质体积呈正相关 , 但是其中的负性成分与消极情绪加工密切相关的右侧杏仁核灰质体积呈负相关 , 这意味着海马灰质体积越大、右侧杏仁核灰质体积越小的个体越有可能具有高自尊 (Wang, Kong, Huang, & Liu, 2016)。贫困的生活会导致个体的海马灰质体积萎缩 , 进而降低记忆和学习功能(Hanson, Chandra, Wolfe, & Pollak, 2011)。基于此的一项最新研究发现 , 贫困和海马灰质体积的关系随个体自尊水平而不同:对于低自尊个体, 生活状况越贫困, 其海马灰质体积越小; 但是对于高自尊个体 , 贫困和海马灰质体积则不存在明显的关系(Wang, Zhang, et al., 2016)。
除了海马和杏仁核 , 自尊与情绪和压力管理相关的脑区 , 如前扣带回 (anterior cingulate cortex, ACC)、右外侧前额叶(lateral prefrontal cortex, LPFC)、左侧下丘脑 (hypothalamus)、右颞顶联合区(temporo-parietal junction, TPJ)的灰质体积也相关。相对于低自尊个体 , 高自尊个体上述脑区的灰质体积较大 , 说明自尊为情绪和压力管理提供了资源 (Agroskin, Klackl, & Jonas, 2014)。另有研究发现个体的生活满意度与右侧海马旁回 (parahippocampal gyrus, PHG)的灰质体积存在正相关、而与左侧楔前叶 (precuneus, PCu)的灰质体积存在负相关 ; 并且, 自尊可以解释生活满意度和这两个区域灰质体积之间的关系 , 即控制个体自尊水平后 , 生活满意度和其灰质体积不再相关 (Kong et al., 2014)。由于右侧海马旁回在情绪记忆 (Murty, Ritchey, Adcock, & LaBar, 2010)、压力管理(Ulrich-Lai & Herman, 2009)以及疼痛和压力感知(Li et al., 2014)等方面具有重要作用。这些研究在一定程度上解释了为什么高自尊可以帮助个体应对压力, 进而提高生活满意度。
1.2自尊与大脑功能的关联
大脑不同区域的结构和功能并非独立 , 而是以相互联结的网络共同发挥作用。默认网络 (default mode network, DMN)是指在静息状态下功能互相关联的脑区网络 , 包括前扣带回 (ACC)、后扣带回 (posterior cingulate cortex, PCC)、内侧前额叶(medial prefrontal cortex, mPFC)、颞顶联合区 (TPJ)等区域 (Fox et al., 2005; Raichle & Snyder, 2007)。大量研究表明自我加工与默认网络活动联系密切 (Gorgolewski et al., 2014; Qin & Northoff, 2011; Sheline et al., 2009)。更有研究揭示自尊与静息态下的默认网络活动存在相关:在静息状态下 , 高自尊的个体后扣带回 (PCC)、楔前叶(PCu)和双侧顶叶下回 (inferior parietal cortex)的活动强于低自尊个体 ; 相反, 低自尊个体的背内侧前额叶 (dMPFC)的活动强于高自尊个体(Fang et al., 2015 )。
由于内侧前额叶在自我加工中发挥着重要作用(杨帅, 黄希庭 , 傅于玲, 2012; Moran, Macrae, Heatherton, Wyland, & Kelley, 2006), 有一些研究专门考察了内侧前额叶与其它脑区的联结性和自尊的关系。在静息状态下 , 自尊与腹内侧前额叶和双侧海马的功能联结存在正相关 , 进一步揭示了的积极的自传体记忆可能作为高自尊的基础 (Pan et al., 2016)。另有研究分别探讨了特质自尊和状态自尊的神经基础。特质自尊对应于个体持久、稳定的自我评价 , 而状态自尊则是暂时性的自我评价, 会随情境而变化 (Brown, 2014)。通过多模态研究分别测量额叶纹状体 (mPFC-striatum)的结构和功能联结性 , Chavez和 Heatherton (2015)发现特质自尊与内侧前额叶和腹侧纹状体的白质结构完整性相关 , 而状态自尊与内侧前额叶和腹侧纹状体的功能联结性相关。由于腹侧纹状体与情绪和奖赏加工有关 (Heller et al., 2013), 自尊与额叶纹状体环路的相关说明自尊同时涉及自我信息加工、情绪加工以及奖赏加工等多个脑区 , 是多个脑区协同作用的结果。
在上述研究中, 研究者通过直接考察自尊与大脑结构和功能的关系 , 揭示了自尊与大脑相应脑区结构及功能的相关性 , 表明自尊作为一种稳定的人格特质, 具有相对稳定的认知神经基础。
2自尊调节大脑对威胁的反应
在日常生活中, 人们随时可能遇到威胁, 任何违背自我动机或预期的信息都可能对人们构成威胁(Jonas et al., 2014)。自尊适应价值的一个重要表现是自尊能够减缓威胁的负面影响 , 帮助个体更好地应对威胁。自尊的这一功能已经得到大量行为研究的支持 (Leary, Tambor, Terdal, & Downs, 1995; Cavallo, Holmes, Fitzsimons, Murray, & Wood, 2012; Ford & Collins, 2010 ; Brown, 2010)。那么 , 在神经水平上 , 不同自尊水平的人对威胁的反应是否存在差异?对此已有相当数量的研究 , 这些研究涉及的威胁主要有三种:社会威胁、死亡威胁和失败。
2.1自尊调节大脑对社会威胁的反应
社会支持是个体自尊的主要来源之一 , 也是个体自尊得以维持的重要基础 (Cooley, 1992; Leary et al., 1995)。因此, 社会拒绝和排斥构成了日常生活中常见的威胁。目前许多研究探讨了不同自尊水平的人面对社会拒绝时大脑反应的差异 , 尤其是注意和情感相关的脑电或脑区反应。
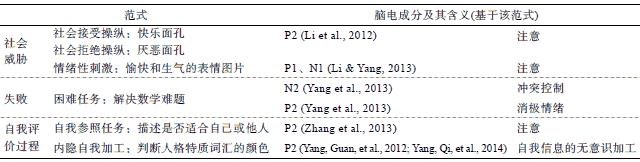
一方面, 自尊会调节大脑对社会威胁的注意反应, 低自尊的个体对社会威胁信息更敏感。从时间进程来看 , 在一项 ERP的研究中 , 研究者向被试呈现快乐和厌恶的面孔分别作为社会接受和拒绝信号 , 结果发现 , 低自尊个体在面对社会拒绝时 P2成分更强 , 这说明低自尊个体为应对威胁分配了更多的注意资源 (Li, Zeigler-Hill, Luo, Yang, & Zhang, 2012)。另一项 ERP研究发现 , 低自尊个体对愉快和生气的表情图片均产生了更大的 P1和 N1成分, 这为低自尊个体对外界情绪的敏感性提供了证据 (Li & Yang, 2013)。从空间结构来看, 一项 fMRI研究同时考察了个体的自尊和注意控制能力 (attentional control), 发现良好的注意控制能力有助于缓解社会拒绝对低自尊个体造成的威胁:当面对社会拒绝时 , 注意控制能力强的低自尊个体比注意控制能力差的低自尊个体产生更强的喙前扣带回 (rACC)反应, 而对高自尊个体, 喙前扣带回的反应不受注意控制能力的影响。由于喙前扣带回与注意管理相关 , 这说明低自尊个体对威胁的敏感性与他们的注意管理能力缺陷有关(Gyurak et al., 2012)。
另一方面, 自尊也会调节个体对社会威胁的情绪活动 , 低自尊个体对社会威胁产生更强的情绪反应。当面对社会拒绝时 , 个体的状态自尊降低, 并且在涉及社会威胁的脑区 , 如与社会拒绝、社会排斥引发的社会疼痛有关的背侧前扣带回 (dACC; Eisenberger, Lieberman, & Williams, 2003;Rotge et al., 2014), 以及与消极情绪加工有关的前脑岛 (insula; Northoff et al., 2006)的激活程度增强 (Eisenberger, Inagaki, Muscatell, Haltom, & Leary, 2011)。但是 , 自尊可以调节个体在社会排斥中的神经活动 , 如在传球(cyberball)任务中 , 低自尊个体对社会排斥感受到更强的社会疼痛 , 他们在背侧前扣带回、腹侧前扣带回、腹内侧前额叶(vMPFC)、脑岛等脑区的反应更强 , 并且他们的背侧前扣带回与内侧前额叶的活动呈正相关 , 而高自尊个体二者的活动呈负相关 (Kashdan et al., 2014; Onoda et al., 2010)。相反, 当面对他人积极的社会评价时 , 低自尊个体在与冲突加工相关的腹侧前扣带回 (vACC)和内侧前额叶 (mPFC)的激活程度更强 , 由于腹侧前扣带回和内侧前额叶涉及对社会评价的情感加工 , 说明低自尊个体对与自我评价冲突的社会评价产生更强烈的情感反应 (Somerville, Kelley, & Heatherton, 2010)。除了注意、情感反应 , 低自尊个体在涉及心智化 (mentalizing)和心理理论(theory of mind)的区域, 如背内侧前额叶 (dMPFC)和后颞上沟 (posterior superior temporal sulcus, pSTS)的激活程度也变高 , 这可能是因为个体在受到社会拒绝后会更多地从他人的角度来思考(Eisenberger et al., 2011)。
2.2 自尊调节大脑对死亡威胁的反应
死亡威胁及应对是社会心理学近 30年来十分活跃的研究领域。根据恐惧管理理论 , 自尊对死亡威胁导致的焦虑具有缓解作用 , 可以降低死亡威胁导致的防御性反应 (Greenberg, Pyszczynski, & Solomon, 1986; Pyszczynski, Greenberg, Solomon, Arndt, & Schimel, 2004)。已有研究表明 , 死亡威胁会导致双侧脑岛、杏仁核以及腹侧前扣带回 (vACC)、喙前扣带回 (rACC)等脑区的特定反应 (Quirin et al., 2012; Shi & Han, 2013)。那么, 自尊是否会调节大脑对死亡威胁的反应呢?在一项 fMRI的研究中 , 研究者向被试呈现与死亡相关、消极但与死亡无关以及中性的句子 , 结果发现 , 个体阅读死亡相关刺激时双侧脑岛的激活程度降低, 而高自尊个体这种反应变化更明显 , 由于脑岛与自我意识加工有关 , 这说明高自尊个体可能通过降低自我死亡意识从而对死亡威胁起到缓解作用; 相反, 低自尊个体的双侧腹外侧前额叶 (VLPFC)和内侧眶额叶皮层 (orbitofrontal cortex, OFC)区域的激活程度变高 , 说明他们对死亡威胁信息的情绪管理需要作出更多的努力 (Klackl, Jonas, & Kronbichler, 2014)。采用词汇分类任务 , 研究者发现高自尊个体应对死亡威胁时杏仁核与腹外侧前额叶 (amygdala-VLPFC)的功能联结增强 , 并且这种功能联结的增强可以降低被试对威胁产生的防御性反应 , 可见自尊通过调节杏仁核与腹外侧前额叶之间的功能联结 , 加强个体对死亡威胁的情绪管理, 从而降低对死亡威胁的防御性 (Yanagisawa, Abe, Kashima, & Nomura, 2016)。
2.3自尊调节大脑对失败的反应
一般而言 , 困难任务通常更容易导致失败 , 威胁个体的自我积极性 , 因此也会对个体构成一种威胁。相关研究分别从时间和空间上表明自尊参与调节大脑对失败的反应。在一项 ERP研究中 , 研究者在被试解决数学任务过程中记录其脑电 , 发现相比于高自尊个体 , 低自尊个体在完成困难的数学任务过程中产生了更强的 P2和 N2效应 (Yang, Zhao, Zhang, & Pruessner, 2013)。由于 P2与消极情绪加工有关 , N2与冲突监控有关 (Folstein & van Petten, 2008), 这说明低自尊的个体在面对困难问题时 , 会体验到更多的消极情绪和冲突。一项 fMRI研究发现, 当启动“聪明”后, 被试如果在当前任务中失败 , 则会减慢对后续任务的反应 , 而启动 “愚蠢”后被试会加快他们的反应, 并且启动“聪明”后犯错误时前扣带旁回 (anterior paracingulate cortex, APC)的反应增强 , 而启动 “愚蠢”后该脑区反应减弱 , 而前脑岛反应增强。由于前扣带旁回与自我概念和行为管理相联系 , 可见, 启动被试积极的自我概念 (如“聪明”)会提高错误监控水平 , 从而促进被试在任务中的自我控制行为(Bengtsson, Dolan, & Passingham, 2011)。
综上所述, 自尊在个体应对威胁的过程中参与调节大脑的反应。从时间进程上看 , 低自尊个体的在早期情绪加工中反应更强(诱发更大的 P1和 N1成分), 他们在面对威胁时会分配更多的注意资源 (诱发更大的 P2成分),体验到更多的认知冲突(诱发更大的 N2成分)。从空间定位上看 , 低自尊个体面对威胁时会产生更强的社会疼痛感 (更强的背侧前扣带回、腹侧前扣带回以及内侧前额叶激活 )和更强的消极情绪 (更强的脑岛、杏仁核激活), 并且他们需要更多的努力来管理情绪 (更强的腹外侧前额叶和眶额叶皮层激活); 而高自尊个体面对威胁能够更好地管理自我意识 (减弱的脑岛活动 )和情绪 (更强的杏仁核 .腹外侧前额叶功能联结)。可见 , 不同自尊水平的个体应对威胁时产生的认知和情感反应存在差异:相比于高自尊个体 , 低自尊个体在认知上对威胁过于敏感 , 在情感上对威胁产生更多的消极体验 , 并且在意识和情绪管理上存在缺陷。可见 , 自尊作为一种稳定的人格特质 , 与情境因素交互影响个体的大脑活动, 进一步影响个体的行为。
3自尊调节自我评价过程中的大脑反应
自尊本质上是个体对自我的评价。不同自尊水平个体的自我评价存在差异 , 因此, 自尊也会影响个体在自我评价过程中的大脑反应。自我评价的研究主要基于两种范式:自我参照 (self-reference)任务和优于平均效应 (better than average, BTA)任务。
自我参照任务通常是让被试判断积极或消极的刺激与自我的相关性 (是否描述自我 )。在此过程中, 人们会倾向于认为积极的刺激更描述自己 , 表现出自我积极偏差 (Cai, Wu, Shi, Gu, & Sedikides, 2016; Epley & Whitchurch, 2008)。研究者通过 ERP和 fMRI技术探讨了不同自尊水平在自我参照任务中的反应差异。 ERP研究发现 , 对低自尊个体而言, 自我积极偏差刺激 (消极.低自我相关 )诱发的 P2潜伏期更长 , 说明他们对自我积极信息分配了更多的注意资源 (Zhang, Guan, Qi, & Yang, 2013)。fMRI研究通过自我参照任务揭示 , 内侧前额叶与自我评价的认知成分更相关 , 而扣带回与自我评价的情感成分更相关 (Moran et al., 2006)。当进行自我评价时 , 低自尊个体对自我相关刺激背侧前扣带回(dACC)反应增强更明显(Yang, Dedovic, Chen, & Zhang, 2012)。而对于高自尊个体而言 , 当呈现积极的自我面孔时 , 高自尊个体在涉及自我相关性的后扣带回 (PCC)和涉及奖赏加工的腹侧被盖区 (ventral tegmental area, VTA)产生更强的反应(Oikawa et al., 2012)。当对自我的积极特质进行评价时 , 高自尊个体的眶额叶皮层 (OFC)激活程度更强 , 说明高自尊个体对自我积极信息产生更强的情感反应(Yang, Xu, Chen, Shi, & Han, 2016)。自尊对自我评价加工的调节作用在内隐层面也有体现。在一项 ERP研究中 , 研究者让被试判断积极和消极的人格特质词的颜色 , 结果发现 , 相比于高自尊个体 , 低自尊个体由自我相关刺激诱发的P2潜伏期更长 (Yang, Guan, Dedovic, Qi, & Zhang, 2012), 进一步研究表明由自我冲突刺激(消极.高自我相关 )诱发的 P2潜伏期更长 (Yang, Qi, & Guan, 2014), 表明他们对违背自我积极性的信息产生了更多的无意识加工。在一项 fMRI研究中 , 研究者采用相同的内隐范式发现 , 对于自我一致性刺激 (积极.高自我相关、消极.低自我相关 ), 高自尊个体在背内侧前额叶和后扣带回产生出了更强的反应(Yang, Dedovic, Guan, Chen, & Qi, 2014)。这些研究说明高自尊个体在自我评价过程中表现出更强的自我积极性。
在优于平均效应任务中 , 被试需要将自己的人格特质与同龄人的平均水平进行比较。基于 BTA的研究发现人们普遍表现出优于平均效应 (Alicke & Govorun, 2005), 反映出人们的自我积极偏差。而当个体受到威胁时 (如消极的社会评价 ),个体倾向于产生更强的 BTA效应(vanDellen, Campbell, Hoyle, & Bradfield, 2011), 并伴随眶额叶皮层(OFC)、内侧前额叶(mPFC)、外侧前额叶 (LPFC)、脑岛和杏仁核活动的增强 (Hughes & Beer, 2013)。进一步证据表明眶额叶皮层的活动根据自尊是否受到威胁而不同:当自尊不受威胁时 , 积极的自我评价会减弱眶额叶皮层的反应及其与颞叶、枕叶和额叶之间的功能联结 , 而当自尊受到威胁时 , 积极的自我评价会增强眶额叶皮层的激活程度, 并且眶额叶皮层与基底神经节(basal ganglia)和额中回 (middle frontal gyrus)的功能联结发生变化 (Beer, 2014)。这说明眶额叶皮层具有自我调节功能 , 当自尊不受威胁时 , 眶额叶发挥着自我监控的功能 , 当自尊受到威胁时 , 个体通过加强自我积极性来抵御威胁。
总之, 自我评价过程涉及前额叶、眶额叶、扣带回等多个脑区 , 但是, 相关脑区的活动受到个体自尊水平的影响 , 高自尊个体在自我评价过程中表现出更强的自我积极性。
4回顾、讨论与展望
4.1回顾与讨论
近年来, 随着社会认知神经科学的迅猛发展 , 涉及自尊的认知神经机制的研究日益增多。本文从三个方面对相关研究进行了梳理:自尊与大脑结构和功能的关联 , 自尊如何调节大脑对威胁的反应, 以及自尊如何调节自我评价过程中的神经活动(具体研究范式以及结果见表 1、表 2)。主要发现有:1)自尊与大脑的海马体积以及杏仁核、前扣带回等脑区的灰质体积存在正相关 , 也与大脑在静息态下的自发性活动 , 以及前额叶与纹状体及海马区域的功能联结存在相关; 2)自尊调节大脑对威胁 (包括社会威胁、死亡威胁和失败 )的反应, 既调节 P1、N1、P2、N2等早期脑电成分 , 又调节前扣带回、内侧前额叶、眶额叶、脑岛等多个脑区活动 , 不同自尊水平的个体应对威胁时的相关脑电和脑区活动存在差异; 3)在自我评价过程中, 内侧前额叶、眶额叶和扣带回等脑区发挥着重要的作用 , 自尊参与调节自我评价过程中的大脑反应 , 既包括脑电反应 (如 P2),也包括后扣带回、腹侧被盖区和背内侧前额叶等脑区的反应。
关于自尊的认知神经研究可以从多个方面加深对自尊的认识和理解。首先 , 这些研究在一定程度上揭示了自尊的神经基础。总体来说 , 自尊是一种复杂的结构 , 既包含对自我的认知又包含对自我的情感体验。因此 , 自尊具有复杂的神经基础, 涉及多个脑区, 包括与自我信息表征和加工相关的脑区 (如前额叶 )、与自传体记忆相关的大脑结构(如海马)、与情绪相关的脑区(如脑岛、杏仁核等)、与奖赏相关的脑区 (如纹状体), 以及相关脑区的功能联结性 (如额叶 .纹状体联结性 )等。同一脑区在不同情境下也会表现出不同的功能 , 比如:与自我评价联系最密切的内侧前额叶 , 在自我评价中更多地涉及认知成分 , 而在社会评价中更多地涉及情感反应。不同脑区之间相互联系 , 相互影响, 共同构成了自尊的神经基础。
其次, 这些研究为自尊的适应价值提供了进一步的证据 , 并且在一定程度上揭示了自尊功能的神经机制。自尊的适应性价值主要表现为高自尊可以帮助个体应对挫折和威胁 , 促进个体心理健康。相应地 , 认知神经视角的自尊研究发现自尊参与调节个体面对威胁时的大脑活动 , 不同自尊水平的个体面对威胁时的大脑活动存在差异。一般而言 , 面对威胁 , 人们会产生近端防御 (proximal)和远端防御(distal)。在近端防御中 , 威胁会立即导致警觉和焦虑 , 行为抑制系统激活 (Behavior Inhibition System, BIS), 个体通过抑制当前威胁相关的信息以避免焦虑 ; 在远端防御中 , 行为趋近系统 (Behavior Approach System, BAS)激活, 此时个体会正视威胁 , 利用心理资源进行自我调节以应对威胁 (Jonas et al., 2014)。和高自尊的个体相比 , 低自尊的个体面对威胁时多采用近端防御, 注意唤醒和情绪反应更强 , 而高自尊个体更多地采用远端防御 , 从而降低威胁带来的负面影响, 更好地应对威胁。此外, 自尊还可以解释相应脑区与生活满意度之间的关系 , 揭示了高自尊的人幸福感更高的原因。这些发现凸显了自尊的重要适应价值:个体的自尊通过影响大脑的神经活动继而帮助个体更好地适应。
4.2研究展望
迄今为止, 尽管自尊的神经研究已经取得了丰硕的成果 , 但是关于自尊 , 依然有大量问题亟待认知神经科学的视角来回答。首先 , 目前的研究虽然揭示了自尊和很多脑区之间的关系 , 但是多数研究只涉及相对单一的脑区以及某些脑区之间的联结性 , 至于这些脑区如何系统、动态地协同发挥作用 , 为自尊相关的复杂行为提供解释 , 无疑是未来研究亟待解决的一个问题。对此 , 研究者曾通过神经模拟的方法提出了基于情绪评价的自尊神经模型 , 描述了特质自尊、状态自尊与情绪加工的神经基础 (Korsten, Fragopanagos, & Taylor, 2007)。该模型认为 , 当外界信息与自我相关时, 信息首先进入长时记忆系统(海马)进行加工, 其结果在一定程度上决定特质自尊 ; 随后, 眶额叶皮层对信息的效价进行加工 , 其结果在一定程度上决定状态自尊 ; 当信息加工为负性时 , 信息进入扣带回 , 此时, 监测到的冲突就会导致个体的情绪反应。但是, 该模型提出时间较早 , 当时相关研究还很有限 , 以至于模型过于简单 , 尚未纳入与情绪加工相关的杏仁核、脑岛 , 与奖赏加工相关的纹状体等诸多重要脑区 , 忽视了自尊神经机制的复杂性 , 更忽视了各个脑区的交互作用。因此 , 该领域亟需更为全面的模型以整合现有的研究。
其次, 由于行为指标和研究方法本身的局限性(如无法对测量对象进行精确区分、无法揭示复杂的内部过程等 ), 许多关于自尊的问题在行为水平上难以明确地回答 , 未来可以利用认知神经指标在时间和空间上的高分辨优势来深入探讨。比如:内隐自尊的神经机制 , 它和外显自尊是否具有不同的神经机制?高自尊和稳定的自尊都对心理健康具有重要的作用 , 二者的神经机制有什么联系和区别?东西方自尊水平的差异到底是源于自我促进动机本身的差异 , 还是由于不同文化下谦虚的重要性存在差异?这些问题在行为层面上争论非常激烈 , 而借助认知神经方法有可能使这些问题得以澄清。例如 , 关于自尊的文化差异是否是因为东方更强调谦虚所导致 (Cai et al., 2011; Kim, Chiu, Peng, Cai, & Tov, 2010), 在行为水平上很难将谦虚和低自尊区分开来。鉴于此 , 有研究采用认知神经科学技术来回答这个问题。一项基于 ERP的自我参照任务发现 , 如果以行为结果 (自我与积极词的联系)作为指标, 则东方文化下的个体自我评价显著低于西方文化下的个体 ; 但是, 如果以表示刺激评价的晚正电位 (late positive potential, LPP)作为指标, 则东西方文化下的个体自我评价差异不显著 (Cai et al., 2016)。和自我报告的行为指标相比 , 脑电指标反映的更多是自动化的自我评价 , 不太受谦虚等情境因素的影响 , 据此推测谦虚可能是导致东方文化下个体自我报告自尊低的主要原因。另一项跨文化的 fMRI研究则进一步为这种可能提供了证据 (Perlmutter, Kuman, & Tssory, 2015)。一般而言 , 在自我评价过程中 , 前额叶的激活程度和自我评价呈正相关 , 而眶额叶则和自我评价呈负相关 , 也就是说 , 前额叶与自我促进有关, 而眶额叶与自我评价的抑制有关。在这项跨文化研究中 , 研究者发现 , 在自我评价过程中 , 东西方文化下的个体的前额叶激活水平类似 , 但是东方文化下个体的眶额叶激活水平显著高于西方文化下的个体 ; 并且, 眶额叶激活程度的差异可以解释东西方个体自我评价的差异。这也表明 , 东西方自尊的差异并非自尊需要本身的差异 , 而是谦虚等其它外在因素影响的结果。
最后, 可以利用认知神经技术探讨个体的自尊是如何形成和发展的。大量关于自尊发展的研究表明 , 自尊源自个体早期的经验和经历 , 会随着发展阶段的不同而不同 ; 在整个发展历程中 , 自尊既表现出了较高的稳定性 , 也表现出了相当的可塑性 (Kuster & Orth, 2013; Orth & Robins, 2014)。研究已经表明自尊和大脑的某些脑区结构及其功能联结性密切相关 , 那么, 大脑的发育和个体自尊的形成与发展存在怎样的对应关系呢?利用认知神经技术 , 我们可以回答许多行为研究无法回答的问题 , 比如:自尊的发展规律是否能在相应的神经指标上得到印证?自尊与对应的大脑结构及功能联结性是如何在个体的发展中逐渐建立联系的?相关脑区的发生、发展规律及其功能的可塑性如何?发展过程中个人的生活经历、经验扮演着怎样的角色并如何反应到神经结构和活动上?已有研究发现回忆童年的自传体记忆时海马和纹状体同时激活 (Oba, Noriuchi, Atomi, Moriguchi, & Kikuchi, 2015),而二者均和自尊密切相关 , 这说明积极的童年记忆可能是高自尊发展的基础。另有研究发现 , 在社会评价情境下 , 相比于儿童和成人 , 青少年的自我意识情绪更强 , 并且伴随着内侧前额叶和额叶纹状体的联结性变化(Somerville et al., 2013)。青少年大脑活动的这种特殊变化也许可以解释青少年自尊的波动性 (Trzesniewski, Donnellan, & Robins, 2003)。可见 , 对于这些问题的解答将极大深化对自尊的特性及其发展规律的认识 , 也将为培养、提升积极健康的自尊提供坚实的基础和指导。
总之, 自尊的认知神经机制研究虽然是近 10年才兴起的一个新的领域 , 但已经积累了相当数量的研究。这些研究在一定程度上揭示了自尊及其作用的神经机制 , 为行为水平上的诸多发现提供了神经证据 , 深化了对自尊的认识。未来的研究可以运用认知神经科学的方法进一步探讨自尊的神经基础 , 特别是针对许多行为水平上无法解决的争议 , 并探索自尊发展的神经机制 , 为自尊的促进和干预提供科学依据。
www.psychspace.com心理学空间网
