
心理科学进展 2008,16(2):250~254Advances in Psychological Science250
崔 芳 南 云 罗跃嘉(北京师范大学认知神经科学与学习国家重点实验室,北京 100875)
摘 要 共情是指个体对他人情绪和想法的感受及理解,它在人类个体和社会的发展中起到了重要作用。近年来,脑成像技术为研究共情的神经基础提供了有力的技术支持,发现了共情的关键功能脑区。基础与临床的实验证据表明,更好地理解共情的神经机制对于孤独症、反社会人格等精神障碍有重要的意义。该文对相关的研究进行了综述并对未来可能的研究方向提出了展望。关键词共情,神经基础,脑成像,精神障碍。
分类号 B8421
什么是共情
1.1 共情的定义
共情(Empathy)指的是对他人情绪和精神状态的理解及对其行为的推测,也就是我们常说的“感同身受”。Singer 提出共情是指这样的能力:1)产生共情的人处于一种情绪状态下;2)这种情绪与另一个人的情绪是同形的;3)这种情绪是通过观察或者模仿另一个人的情绪而产生的;4)产生共情的人能意识到自己当前情绪产生的原因在于他人而非自身[1]。
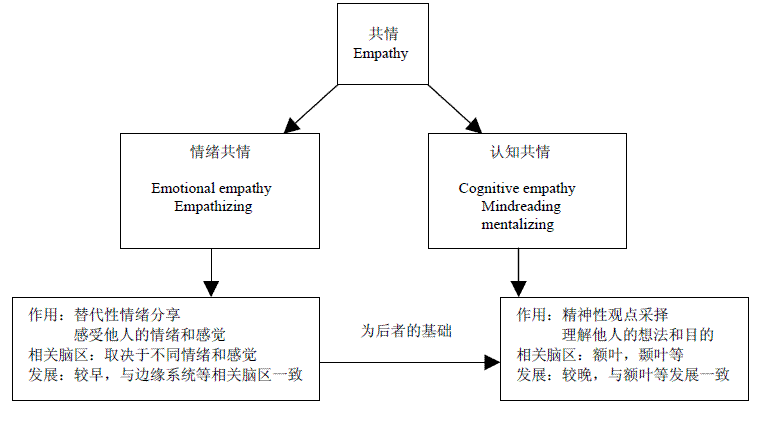
对共情的认识有多种不同的看法。其中Gladstein于1983年提出的两成分理论影响最大。这种理论认为共情包括认知共情和情绪共情,其中前者指的是对他人目的、企图、信仰的理解;后者指对他人情绪状态的感受[2]。
对共情的定义及划分存在着很多不同看法。产生这种现象的原因一方面是共情本身的复杂性;另一方面则是研究者的出发点不同。相信这种争议在短期内很难结束。只有随着研究的继续深入去寻求更加贴切,更加本质的理论解释。
1.2 情绪共情和认知共情
从共情的功能上来看,可以划分为情绪共情和认知共情。
情绪共情(Emotional Empathy)指的是对他人情绪的一种替代性分享。脑成像实验结果表明,他人的情绪可以激发个体本身与这种情绪或感觉相关脑区的活动,将他人的情感表征转化成个体自身的情感表征,从而令个体对他人的情绪“感同身受”。因此,情绪共情的神经活动与诱发共情的情绪或感觉的性质有关。
情绪共情在个体的发展中出现比较早,研究表明对他人情绪的感受很早就出现了,是个体进行社会性交往的第一个工具。14个月大的婴儿会对其他婴儿的哭泣做出类似的哭泣反应[3]。情绪共情出现的时间和与情绪相关脑区,比如边缘系统,杏仁核等的发育是一致的。
除了感受和理解他人的情绪以外,人类还有一项独特的能力—— “ 认知共情”( CognitiveEmpathy),即对他人的想法,意图进行理解,并由此推测其未来的行为。认知共情的发展相比于情绪共情出现较晚,采用“失言”任务(给被试阅读一个失言的故事,问被试几个问题,比如:故事里有人说了不应该说的话吗?谁说了?为什么不应该说?这项任务要求被试具有一定的认知共情能力。)进行测试发现:在3 到4 岁时,儿童开始能够了解到他人具有与自己不同的情绪和想法。到了6 岁,儿童的观点采择能力初步形成,到了11 岁左右,儿童才具备了比较完善的观点采择能力[4]。这一点和与高级认知相关的顶额叶脑区发展是相一致的。情绪共情和认知共情相互独立,又紧密联系。二者的有机结合增强了个体的社会交往能力。情绪共情可以压制暴力行为,产生利他行为的动机,认知共情则可以帮助我们选择最合适的办法去帮助别人[5]。两者的融合在社会环境下是非常重要的,某一方面的失调会造成社会交往的障碍或问题。
1.3 共情的生物学意义
一般认为,共情可以让社会成员之间更容易形成关系,从而促进了利他行为的产生并抑制了反社会行为。从达尔文的生物进化观点来看,共情对人类的生存和人类社会的形成及发展有着重要的意义。共情的生物学意义主要有以下两点。
首先是认识论上的功能。通过分享他人的情绪,我们可以理解他人的感受。共情令我们更加准确和迅速地判断他人的行动,同时也提供了重要的环境信息。它就像一台高效的计算机,帮助我们从周围世界获取有价值的信息[6]。
其次是社会性功能。研究表明:有共情障碍的个体常常出现交往困难,甚至反社会行为。共情能力可以增进社会交流水平和社会凝聚力。对于发展道德和抑制暴力有重要意义[6]。
共情是生物进化的产物。是人类生存的选择。虽然经过了生物进化的不断完善,人类的共情还是有其局限性的。很多时候人们都是在自身经验的基础上去理解他人的感受和意图。这常常与他人的实际情绪或想法有所差距。大量社会心理学研究表明,个体在对他人观点进行采择时会经常性犯无意识的错误。有假设认为:这种错误理解是基于一种将自身观点赋予他人的自动化倾向[7]。这种效应被称为“共享表征的自我中心假设”(the egocentricassumption of shared perspectives)[8]。这种从自身角度出发的共情常常造成很多社会误解。
2 共情的神经基础
从当前的认知神经科学研究结果来看,共情的神经基础主要由以下几部分构成:动作知觉和情绪分享系统,主要作用是诱发共情发生的主体与被观察客体产生相似体验;自我–他人意识系统,这部分使得主体能明确地意识到自己情绪产生的根源是他人而非自身;另一个重要部分就是由额叶起主导作用的精神调控系统,其作用是整体调节和意识控制。这三部分的有机结合和共同作用使得共情成为一个比较完善的社会工具,对人类生存和人类社会的发展起到了关键作用。下面对这几个部分的研究成果进行详细介绍。
2.1 知觉–动作模型与情绪分享
个体与他人之间的“表征共享”是共情的基础[9,10]。所谓“表征共享”就是当个体知觉到他人的动作或者情感的时候,其大脑中表征相应动作或情感的部位会被激活,从而令个体产生同形的表征。这种能力似乎是人类与生俱来的,新生儿自发动作为其与外界的自然接触提供了充分条件[11]。知觉–动作模型由Preston 和de Waal 于2002 年提出。他们指出共情是物种进化的产物,类似的模仿能力在猿、老鼠等非人类的社会性哺乳动物中也有发现,它使得个体对危险环境的学习能力得到了提高并且促进了同类之间亲社会行为的发生。对于人类特有的共情能力来说,其基本的假设是:当个体知觉到他人的行为时会自动激发其与该行为有关共情Empathy情绪共情Emotional empathyEmpathizing认知共情Cognitive empathyMindreadingmentalizing作用:替代性情绪分享感受他人的情绪和感觉相关脑区:取决于不同情绪和感觉发展:较早,与边缘系统等相关脑区一致作用:精神性观点采择理解他人的想法和目的相关脑区:额叶,颞叶等发展:较晚,与额叶等发展一致为后者的基础
的个人经验的表征[12]。
镜像神经元(mirror neuron)的发现为动作–知觉模型提供了有力的证明。Rizzolatti 与Gallese 在恒河猴的大脑腹侧运动前皮质上发现了这种神经元。它们在恒河猴进行目的性动作时或者当恒河猴观察到另一个同类做出相同动作时都被激活[13]。最近有研究发现,当一个动作最后的部分被仅仅暗示出来的时候,镜像神经元也会被激活。这说明这部分特定神经元是对动作的表征做出反应而不是对动作本身反应[14]。镜像神经元的存在证明了在观察到他人进行某种活动和自身做出该活动时会激活相似的脑区。这为人类共情的发生提供了生物基础[15]。举个例子:如果看到他人悲伤和自身悲伤激活了同样的脑区,那么在观察到他人悲伤的表情时,个体自身的悲伤情绪也在一定程度上被诱发了,这样,“共情”就发生了。
Jackson 等人在一项实验中给被试出示处于危险状态的肢体图片(比如一根夹在门缝处的手指)。肢体部分经过模糊处理,消除了年龄和肤色的干扰。在指导语中令被试分别想象这根手指是自己的/他人的/假肢。然后在疼痛程度量表上进行评定。脑成像结果发现:当被试想象手指是自己的和想像手指是他人的条件下,与疼痛相关的脑区都被部分激活了。而在想象手指是假肢的条件下,这部分脑区则没有被激活[16]。这说明,个体自身和他人的相同感受在大脑中的加工有重叠部分,也支持了共情与相应的情绪有共享的神经回路这一观点。
在厌恶情绪的研究中也发现了类似的结果。有一项研究将被试观察他人厌恶表情的面孔时大脑活动与被试闻到恶心气味气体时的脑部活动采用fMRI 技术做了对比。发现两种条件都激活了后脑岛和扣带前回的相似部位,而这两个部位在先前的研究中都被发现与厌恶情绪有关,而脑岛则是厌恶情绪的关键脑区[17]。这说明共情发生时,相同情感或感觉在大脑中被诱发了。
共情诱发相关脑区的程度和部位受到多种因素的影响。比如:产生共情的主体和被观察的对象之间的关系。Singer 在2004 年的一项实验中令被试观察自己的爱人受到疼痛刺激,之后令被试观察陌生人受到疼痛刺激,通过两者的对比得出了在疼痛共情中只有与情感性质相关的部分会调节共情,而与感觉性质相关的部分则对共情没有影响的结论[18]。在Singer 的另一项实验中,第一阶段利用一个“监狱两难游戏”(Prisoner’s Dilemma game)来诱发被试对两名助试不同的情感态度:满意或不满意。第二阶段对两名助试进行疼痛刺激,采用fMRI 记录被试的大脑活动情况。研究发现不论男性还是女性被试看到自己满意的助试受到痛苦刺激时都产生了共情。但看到自己不满意的助试遭受痛苦刺激时,男性被试共情不明显,女性被试的共情有所减弱[19]。
2.2 自我–他人意识
共情的一个重要特征是发生共情的个体虽然与他人发生了相似的情感,但明确地知道他人才是自己这种情绪发生的根源。也就是说,对自我和他人的有明确区分。
在个体发展中,自我意识的形成是共情能力成长的关键[20]。自我意识并不是依赖于一个特定的脑区,而是大脑各个部分交互作用的结果。在自我意识中,大脑右半球起了主导作用[21]。Keenan 发现病人在右半球麻醉的时候会对自己的面孔和他人面孔区别灵敏度下降[22]。在采用“自我参照范式”对自我–他人意识进行研究的脑成像实验中发现:与他人参照加工相比,自我参照加工计划激活了内侧前额叶[23,24]。Craik 等在1999 年的一项PET 研究中也发现了自我意识加工显著激活了右侧额叶,认为自我概念的表征定位于右侧额叶[25]。Zysset 等发现自我参照加工激活了额下回,内侧前额叶和扣带回[26]。从近期的研究结果来看:几乎所有与自我意识相关的任务都激活了内侧前额叶(MPFC)和扣带回。
2.3 精神适应性与自我调节
我们周围的环境时刻充满了各种刺激,如果共情仅仅按照知觉–动作模型发生作用,那么每个人都会一直被他人的情绪所影响。因此,在共情系统中除了发生情感分享的知觉–动作子系统和区别自我和他人的子系统之外,还有一个提供精神适应性和起情绪调控作用的系统,从而使得共情成为一个较为理想的社会交流形式。
起这一作用的脑区主要是顶额叶脑区。前额叶在情绪控制过程中起了基本的作用[27]。研究发现双侧前额叶损伤的病人在对他人观点采择(perspective-taking,指个体把自己的观点与他人的观点区分开并协调起来的能力)的能力测验中表现出了明显障碍[28]。在一项研究额叶损伤病人视觉观点采择和欺骗能力的实验中,研究者发现右侧额叶对观点采择的能力有更大的影响[29]。Ruby 与Decety 进行的一项PET 实验中采用了动作假想范式(motor imageryparadigm),令被试观察做出不同动作的身体图片,通过指导语令被试想象做出该动作的是他本人(第一人称)或者做出该动作的是另一个个体(第三人第16 卷第2 期 共情的认知神经研究回顾 -253-称)。比较两种条件下大脑活动的差异。分别在第一人称和第三人称状态下对其脑部活动进行扫描的结果发现前额叶皮层中部,后部扣带回和额极在观点采择中起重要作用[30]。
3 共情障碍与精神疾病
大多数共情障碍被认为是额叶,特别是前额叶受损的结果。通过本文对共情相关脑区的总结看来,很多脑区受到损伤都有可能造成共情障碍。共情障碍也应表现为多种不同的形式。
根据共情的社会功能,它可以帮助人们理解和感受他人的情绪和心理状态,从而促进人们之间的相互关系的建立和发展以及亲社会行为的发生。那么反社会人格障碍(Antisocial personality disorder)与共情是否有关系呢?Adam Smith 于2006 年提出反社会人格障碍患者是情绪共情低下同时认知共情发达,共情缺陷所造成的对他人情绪状态特别是对恐惧和悲伤感受能力的下降是形成反社会人格的一个主要原因[5]。
孤独症也是共情研究者们关注的一种精神障碍。孤独症患者一般表现出广泛的社会交往缺陷。他们不愿意和别人接触,好像对他人的感受也无动于衷。一般认为孤独症是一种共情缺陷的症状[31]。但关于这种共情缺陷的神经基础却有不同的意见。一种观点认为孤独症仅仅是对他人观点采择能力缺陷的表现,其情绪共情能力是正常的[8]。另一种看法认为孤独症的患者不仅仅是对他人想法和意图理解能力的缺陷,同时对他人情绪感受的理解也有缺陷,他们在社会交往中是笨拙而冷漠的[32]。从行为和认知神经的研究结果来看,近期的大部分研究证明了前者。关于孤独症的成因和内在神经机制还需要借助现代脑成像技术进行进一步研究。
有研究者[5]认为以思维障碍、情感淡漠、出现妄想等特点为症状的精神分裂人格障碍(schizoidpersonality disorder)患者是情绪共情和认知共情受损的结果。
威廉姆丝综合症(Williams syndrome)的患者表现出对陌生人过度友好,在新环境中不感到害羞,但对新情景表现出极度焦虑,对他人情感非常容易发生共情,是一种情绪共情过度发达,认知共情不足的情况[5]。
这些精神疾病都被认为与共情缺陷有关,但其具体的关系和相关的神经基础仍不明。
由此可见,对共情神经基础的研究会进一步加深对患有社会性认知障碍人群的理解。同样,对特定的精神疾病的研究也会为共情神经机制的发现提供重要线索。
4 未来研究展望
共情涉及社会心理学、发展心理学、认知神经心理学、比较心理学等多个学科分支,是一个比较复杂研究课题。其研究还处于初级阶段,尚未形成成熟的理论。还有很多有待解决的问题。
不同情绪(快乐,悲伤,厌恶,恐惧等)产生时激活的脑区有待进一步研究,相应情绪发生共情时激活的脑区也未明确。顶额叶在共情中是如何调节控制情绪的?情绪共情和认知共情是否在神经基础上相互独立?都是当前研究的问题。
另外,Singer 的一项研究表明,男性对缺乏好感的对象共情下降明显,而女性则不明显。从日常经验来看,女性似乎更容易产生共情,产生共情的对象更广泛[19]。那么,共情是否存在性别差异,这种差异存在怎样的神经基础呢?
共情与发生共情的个体自身情绪的相互关系是怎样的?对同一种外部情绪,个体自身情绪状态不同时发生共情所激活的脑区是否有不同?如果有,是怎样的不同?如何去解释这种不同。如何在自我–他人意识(self-other awareness)和不同情绪(边缘系统,杏仁核等)的神经基础上探索这一问题?共情在心理治疗方面是一个重要的概念。如何通过发掘共情的神经机制而发现孤独症、反社会人格障碍等精神疾病的内在机制从而更好的治疗?已经有很多研究者将共情与一些精神失调联系起来,并试图用共情缺陷来解释这些疾病的成因。这方面的研究需要有现代脑成像技术的支持,从而发现其神经结构上的确切联系。
总之,共情的研究,特别是其认知神经方面的研究才刚刚起步,需要研究者应用现代技术和新的方法深入探讨。
参考文献
1 Singer T. The neuronal basis and ontogeny of empathy andmind reading: review of literature and implications for futureresearch. Neuroscience and Biobehavioral Reviews, 2006, 30:855~863
2 Gladstein G A. Understanding empathy: Integrating counseling,developmental, and social psychology perspectives. JournalCounseling Psychology, 1983, 30: 467~482
3 Meltzoff A N, Moore M K. Newborn infants imitate adult facialgestures. Child Development, 1983, 54: 702~709
4 Stone V E, Baron-Cohen S, Knight R T. Frontal lobecontribution to theory of mind. Journal of CognitiveNeuroscience, 1998, 10: 640~646-254- 心理科学进展 2008 年
5 Smith A. Cognitive empathy and emotional empathy in humanbehavior and revolution. The Psychological Record, 2006, 56:3~21
6 Decety J, Jackson P L. The functional architecture of humanempathy. Behavioral and Cognitive Neuroscience Reviews,2004, 3(2): 71~100
7 Green J D, Sedikides C. When do self-schemas shape socialperception? the role of descriptive ambiguity. Motivation andEmotion, 2001, 25(1): 67~83
8 Fenigstein A, Abrams D. Self-attention and the egocentricassumption of shared perspectives. Journal of ExperimentalPsychology, 1993, 29: 287~303
9 Decety J, Sommerville J. A Shared representation between selfand others: a social cognitive neuroscience view. Trends inCognitive Science, 2003, 7: 527~533
10 Jeannerod M. To act or not to act: Perspective on therepresentation of actions. Quarterly Journal of ExperimentalPsychology, 1999, 52A: 1~29
11 Decety J. Naturaliser l’empathie [Empathy naturalized].L’Encéphale, 2002, 28: 9~20
12 Preston S, D de Waal. Empathy: Its ultimate and proximatebases. Behavioral Brain Science, 2002, 25: 1~72
13 Rizzolatti G, Fadiga L, Gallese V, et al. Premotor cortex andthe recognition of motor actions. Cognitive Brain Research,1996, 3(2): 131~141
14 Umilta MA, Kohler E, Gallese V, Fogassi L, Fadiga L, et al. Iknow what you are doing: a neurophysiological study. Neuron,2001, 31:155~165
15 Jackson P L, Meltzoff A N, et al. Neural circuits involved inimitation and perspective-taking. NeuroImage, 2006, 31:429~439
16 Jackson P L, Rainville P, Decety J. To what extent do we sharethe pain of others? Insight from the neural bases of painempathy. Pain, 2006, 125: 5~9
17 Wicker B, Keysers C, Plailly J, et al. Bothe of us disgusted inmy insula: the common neural basis of seeing and feelingdisgust. Neuron, 2003, 30: 655~664
18 Singer T, Ben S, O’Doherty J, Holger Kaube, Raymond JDolan, Chris D, Frith. Empathy for pain involves the affectivebut not sensory components of pain. Science, 2004, 303:1157~1162
19 Singer T, Seymour B, Doherty J P, Stephan K E, Dolan R J,Firth C D. Empathic neural responses are modulated by theperceived fairness of others. Nature, 2006, 26: 466~469
20 Lewis M. Social cognition and the self. Early social cognition:understanding others in the first months of life. Mahwah, NJ:Lawrence Erlbaum Associates Inc, 1999. 88~98
21 Keenan J P, Gallup G G, Jr, Fslk D. The face in the mirror: thesearch for the origins of consciousness. New York, NY:HarperCollins/Ecco, 2003. 278
22 Keenan J P, Nelson A, O’Connor M, Pascual-Leone A.Self-recognition and the right hemisphere. Nature, 2001,305~409
23 Northoff G, Bermpohl F. Cortical midline structures and theself. Trends in Cognitive Science, 2004, 8 (3): 102~107
24 Ying Z. Neuroimaging studies of self-reflection. Progress inNatural Science, 2004, 14(4): 296~302
25 Craik F I M, Moroz T M, Moscovitch M, et al. In search of theself: a positron emission tomography study. PsychologicalScience, 1999, 10: 26~34
26 Zysset S, Huber O, Ferstl E, et al. The anterior frontomediancortex and evaluative judgment: an fMRI study. Neuroimage,2002, 15: 983~991
27 Grimm S, Schmidt CF, et al. Segregated neural representationof distinct emotion dimensions in the prefrontal cortex.Neuroimage, 2006, 30: 325~340
28 Rowe A D, Bullock P R, Polkey C E, Morris R G. Theory ofmind impairments and their relationship to executivefunctioning following frontal lobe excisions. Brain, 2001, 124:600~616
29 Stuss D T, Gallup G, Alexander M P. The frontal lobes arenecessary for theory of mind. Brain, 2001, 124: 279~286
30 Ruby P, Decety J. What you believe versus what you thinkthey believe: a neuroimaging study of conceptual perspectivetaking. European Journal of Neuroscience, 2003, 17:2475~2480
31 Baron-Cohen S,Belmonte M K. Autism: a window onto thedevelopment of the social and the analytic brain. AnnualReviews Neuroscience, 2005, 28: 109~126
32 Baron-Cohen S. The extreme male brain theory of autism.Trends in Cognitive Science, 2002, 6: 248~254
A Review of Cognitive Neuroscience Studies on EmpathyCUI Fang, NAN Yun, LUO Yue-Jia
(State Key Laboratory of Cognitive Neuroscience and Learning, Beijing Normal University, Beijing 100875, China)Abstract: Empathy is the understandings and feelings about other people’s emotion and thoughts, which is vital forindividual development and human society. Recently, the neural basis of empathy is attracting more and moreattention in neuroscience research. Functional imaging studies have demonstrated activation of some key areasassociated with empathy in the human brain. Basic and clinical evidence also suggests that a better understanding ofneural basis of empathy has important implications for autism and anti-social personality. This article reviewsempathy-related research and highlights its possible future.Key words: empathy, neural basis, brain imaging, mental disorders.
www.psychspace.com心理学空间网
